![]()
|
|
|
|
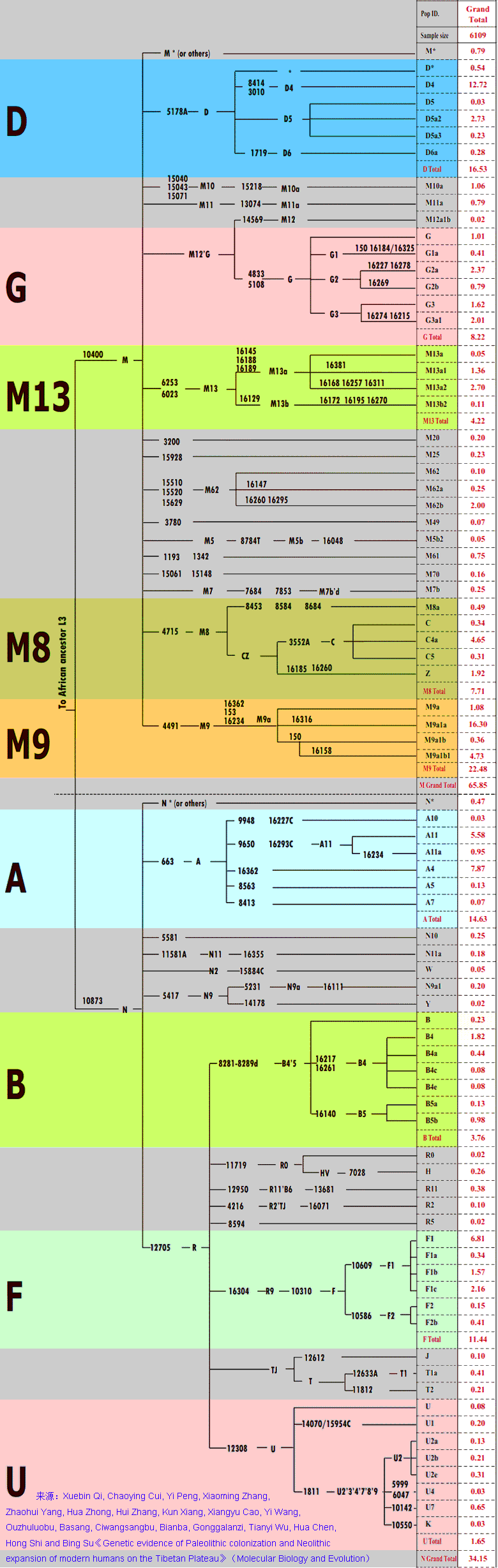
原数据文章出处:Genetic polymorphism investigation of the Chinese Yi minority using PowerPlex® Y23 STR amplification system。 单倍群分析:分子人类学论坛.Ryan。当中M117的73例,主要是CTS1642占31例(42.5%),再者是上游的F5(19.1%)及F5970(20.5%);O1a主要是F140,F492少见。 凉山彝族自治区是全国最大的彝族聚居区,这份单倍群数据像个大杂烩,略估有藏川滇一带起源的D1及西南土著F2、黄河上游的N2、源自广西的O2a-M95及源自华东的O1(如明显东部特色的F140);O3系统有早期分出的9例M324、有西北来源的F155支,F444支也不少,相对002611只有少量。O3中最大头是M117,占23.5%,当中又以CTS1642占多数,占10%,主要是B456支。因为Y7080是F14523的下游分支,B456已分析设定,所以只有用F14523*这样的表达才会包含有Y7080,Oα1c1b4*应属B456这一支的,与Y7080无关,CTS1642七例也应不包含有Y7080。 可以认为CTS1642-B456等(xY7080、CTS9713)及O3的早期分支M122、M324、P201等是彝族的源头,早期应于湘西向西分出,而D1、F2是西迁过程融入的西南土著。其他算是从周边进来的流民,当中,M95是融合进来的濮人,O2的源头在广西;O1a系统来自华东远方;N2及F5的F155分支来自西北;F444分支来自中原东部;002611可能来自楚人。 从凉山彝族的CTS1642看其下游分支的迁徙及分布:B456等分支从湘西迁西南,并融合成彝族,缅甸缅族的CTS1642(xY7080)应与其有同源关系,彝、缅属同个语族;Y7080支没有直接从湘西走进西南,而是首先南迁岭南,到达珠三角,4千年前左右才伴随着西樵山文化的双肩石器的向西传播走进西南,傣族等当中的CTS1642主要是Y7080支;CTS9713支在湘西原地融合到M95、M7孟苗人群当中并形成荆蛮族群,后来CTS9713随着三苗迁徙山东与东夷混合,被黄帝集团打败后散落到韩国等地,北方人群也有少量CTS9713。 来源:Yali Xue, Tatiana Zerjal, Weidong Bao等. Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times 石家庄152个样本:M134*=15.1%、M117=16.4%、002611=13.2%、M119=8.6%、M217=9.9%、M15=2%、其他=34.9%。(分子人类学论坛.雄镇散人) 在韩国506例当中,O3谱系有224例占44.3%,其中O3*-M122占0.4%、O3a1*-KL2占0.8%、O3a1c-002611占14.2%、O3a2*-P201占8.9%、O3a2b1-M7占0.6%、O3a2c*-P164占0.6%、O3a2c1*-M134占9.5%、O3a2c1a-M117占9.3%。(据sahaliyan)
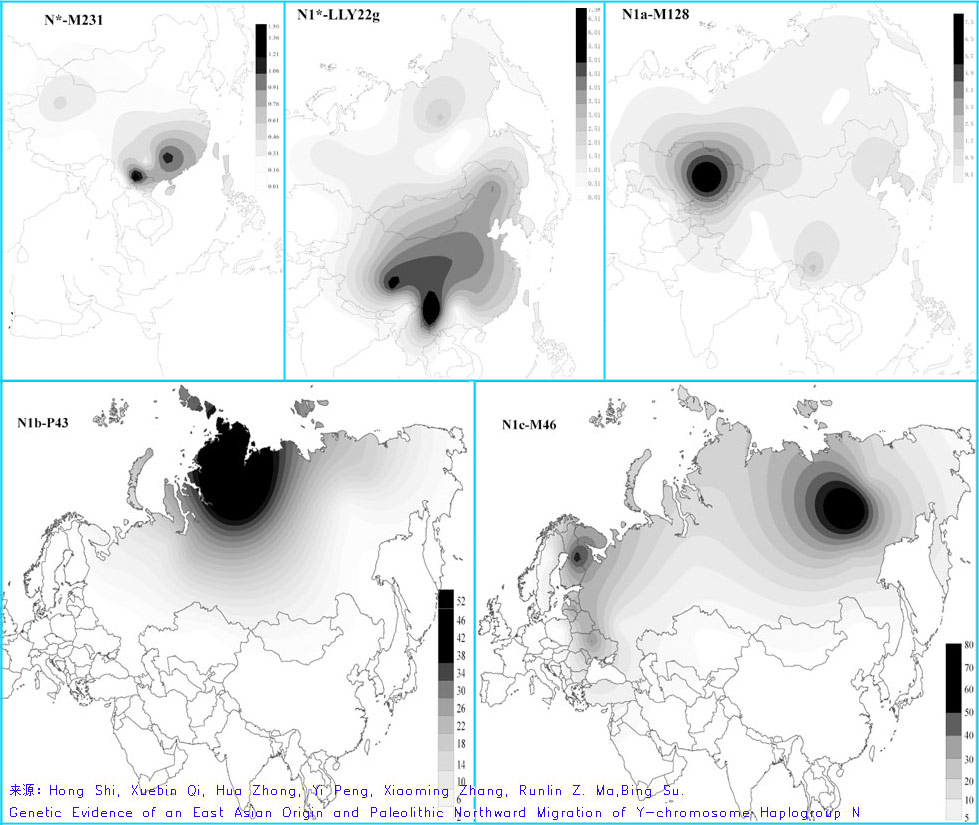
来源:Xuebin Qi, Chaoying Cui, Yi Peng, Xiaoming Zhang, Zhaohui Yang, Hua Zhong, Hui Zhang, Kun Xiang, Xiangyu Cao, Yi Wang, Ouzhuluobu, Basang, Ciwangsangbu, Bianba, Gonggalanzi, Tianyi Wu, Hua Chen, Hong Shi and Bing Su《Genetic evidence of Paleolithic colonization and Neolithic expansion of modern humans on the Tibetan Plateau》(Molecular Biology and Evolution) (sahaliyan的帖子) (注意:N*、O*、O1、O2罕有,说明藏族的O系、O3系统基本不包含云南或南方因素;在O3系统中,M117占到90%,这有点意外,而印度东北的卡西人、加若人M134*比例高于M133,同时还有M122*,看来西南藏缅语族的O3系统还不能简单地看作是汉藏O3系统的南向延伸。) 石宏等《Y单倍群N的东亚起源和旧石器时期的向北迁徙》(2013)中的图表 几句译文:基于N在东南亚的大部份地区缺失,新数据不支持N起源于东南亚(指的是中南半岛)。从几个支系如N1*-LLY22g和N*-M231等未细分类型的分布来看,N更像是起源于中国南部。N*-M231的未细分类型最常见于中国南部的壮侗人群、南方汉族、藏缅语族、苗瑶族等,而完全缺失于西伯利亚、中亚及欧洲。N1*-LLY22g的未细分类型从中国南部到北亚皆有分佈,最高频区见于藏缅族群。N1a-M128的分布与N1*-LLY22g较为类似,但平均来说较为低频,其高频点见于中亚的哈萨克族(8.1%)。最后,N1b-P43及N1c-M46皆富集于西伯利亚,相异的是,N1b-P43在北亚的分布频率是由北往南递减的,N1c-M46则是由东向西递减。Y-STR计算得出的N1a-M128年龄仅有3750年左右,而中亚哈萨克族的N1a-M128不像是中亚本地起源,更像是源自东亚的扩张。(据natsuya)
|
{kind=link}