![]()
|
|
|
|
从分子人类学图表分析日本人的起源 地学演绎 莫如波 2010.2.15
下面的图表源自日本人的网站,转自分子人类学论坛natsuya网友的帖子。
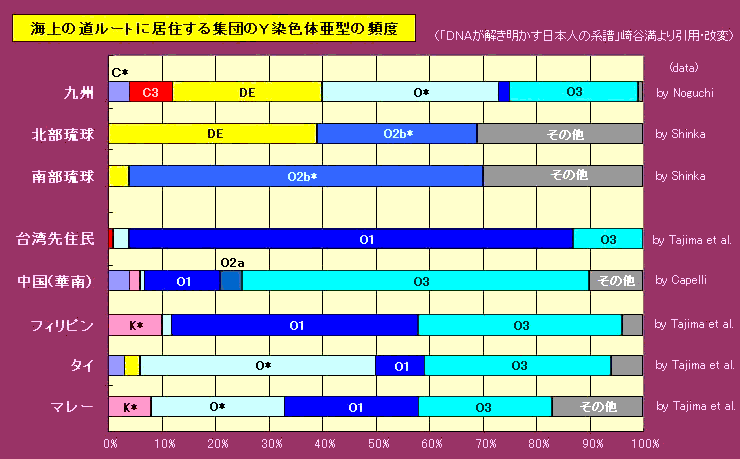
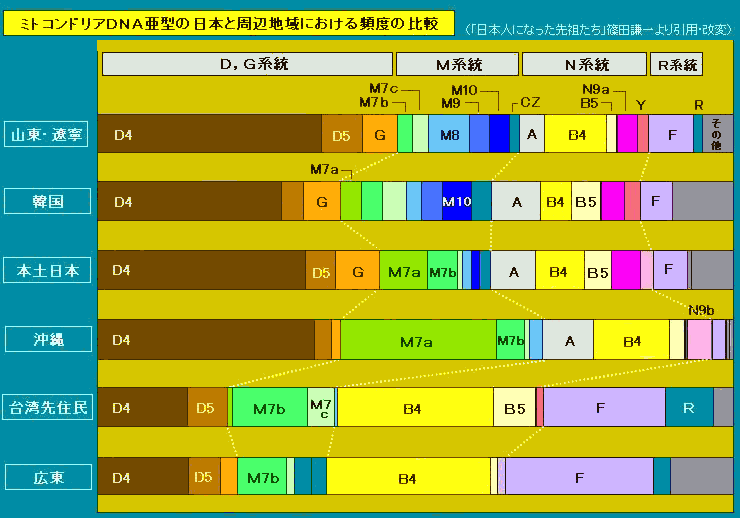
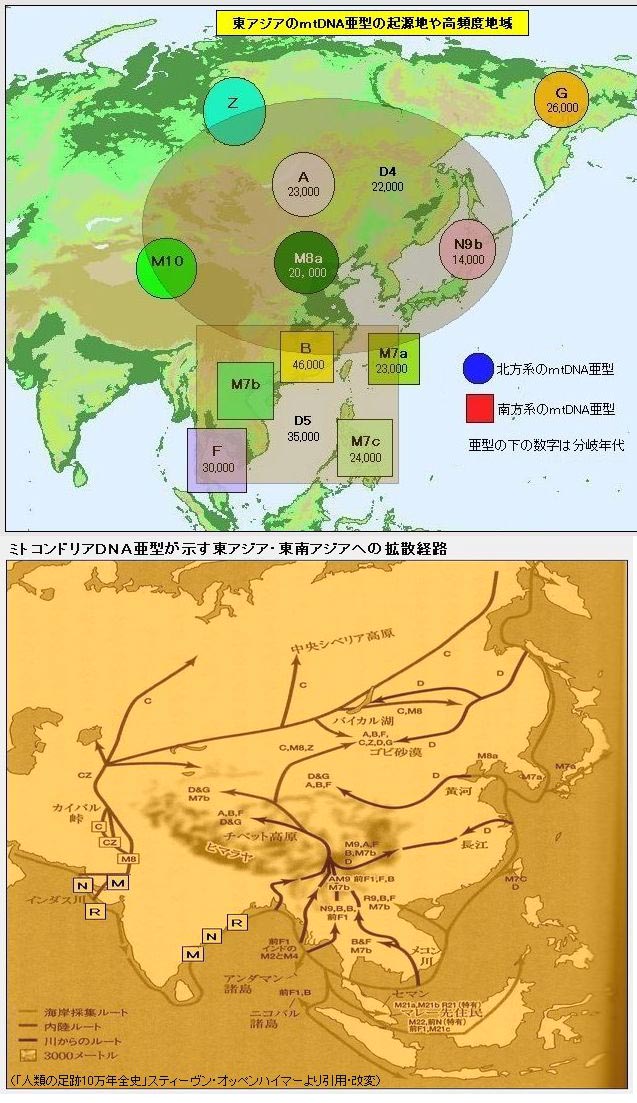
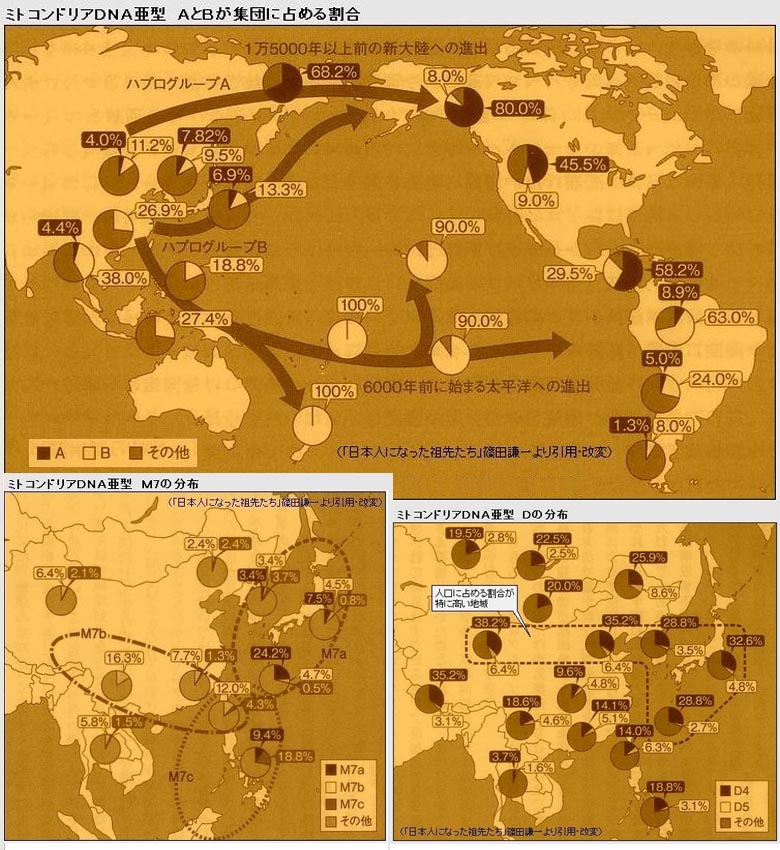
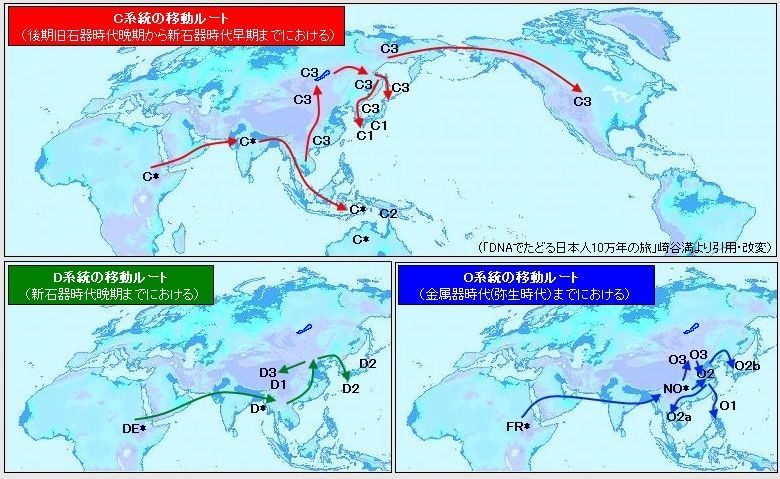
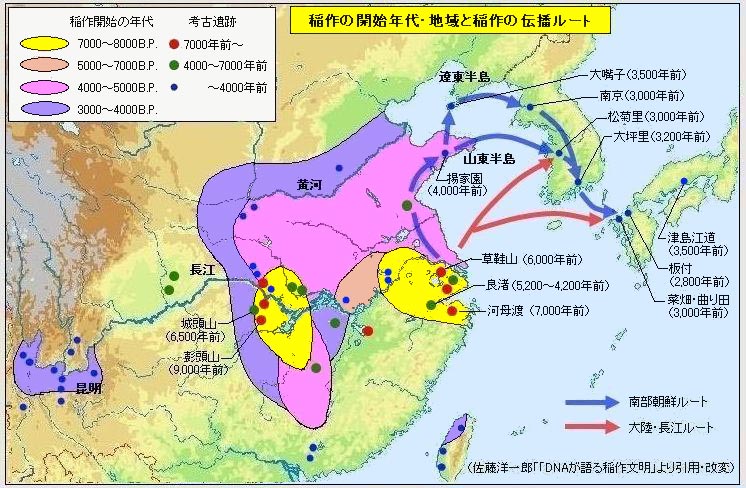
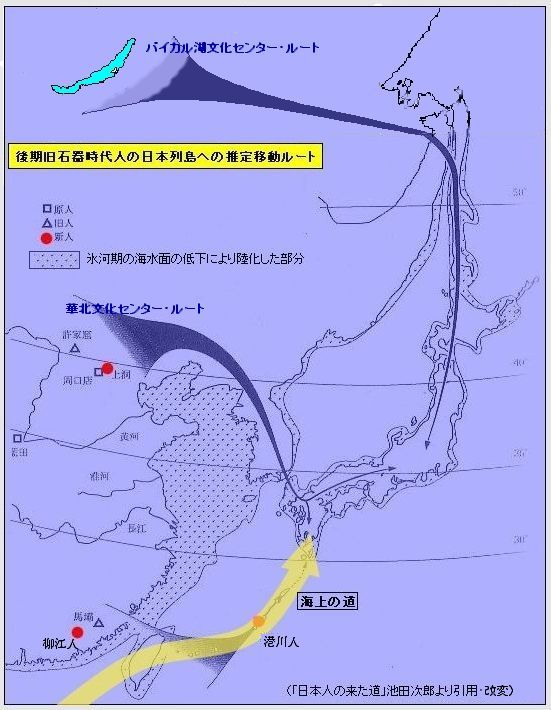
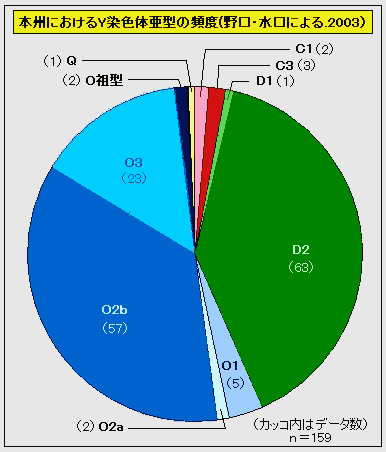
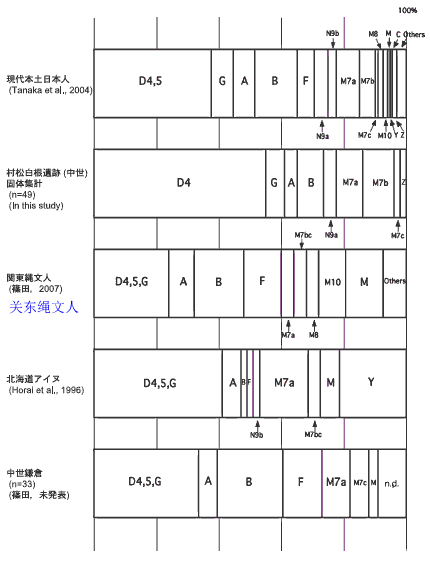
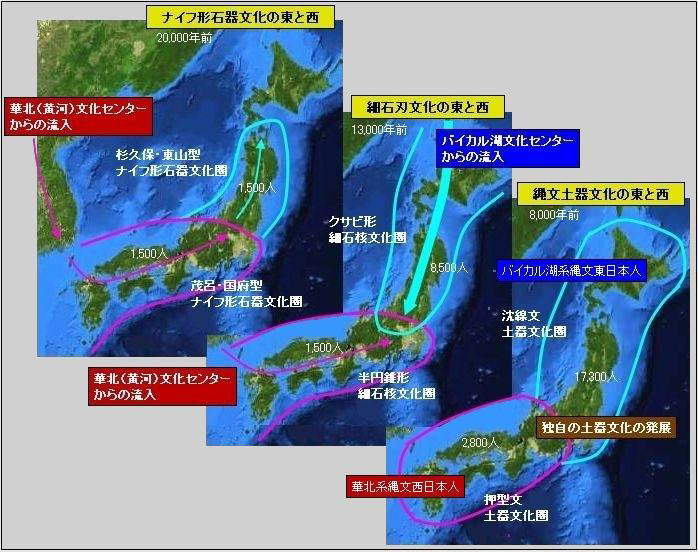
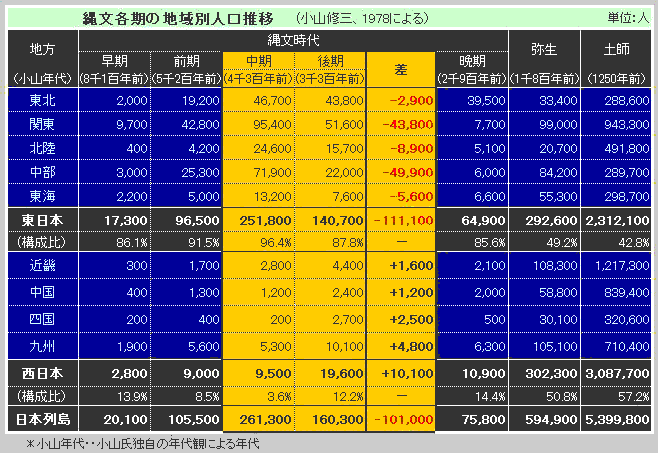
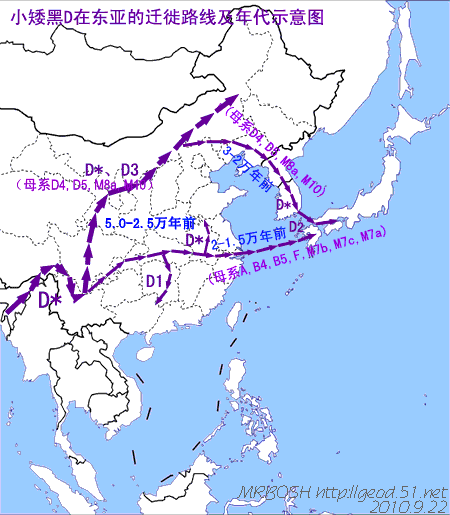
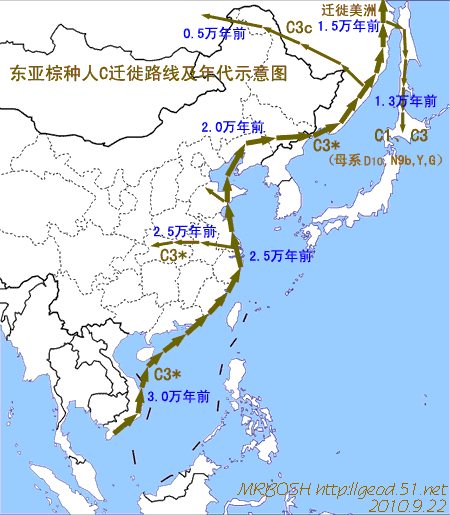
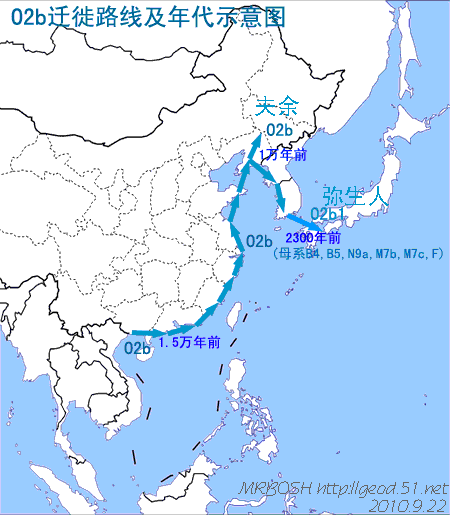
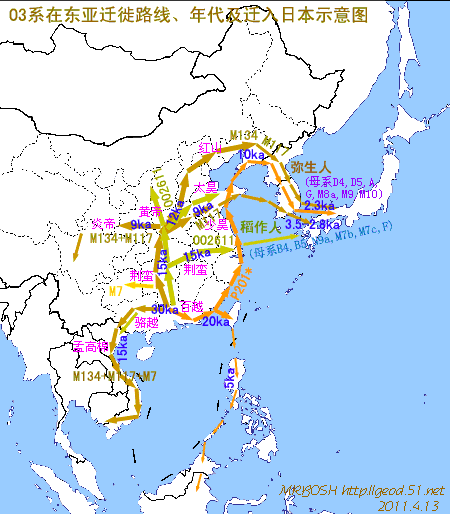
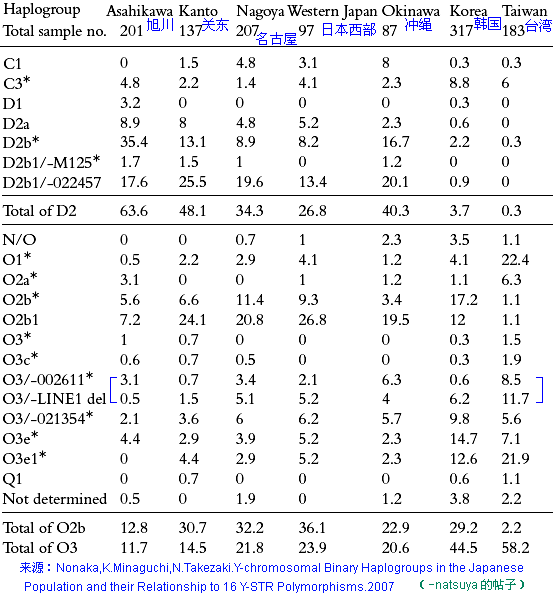
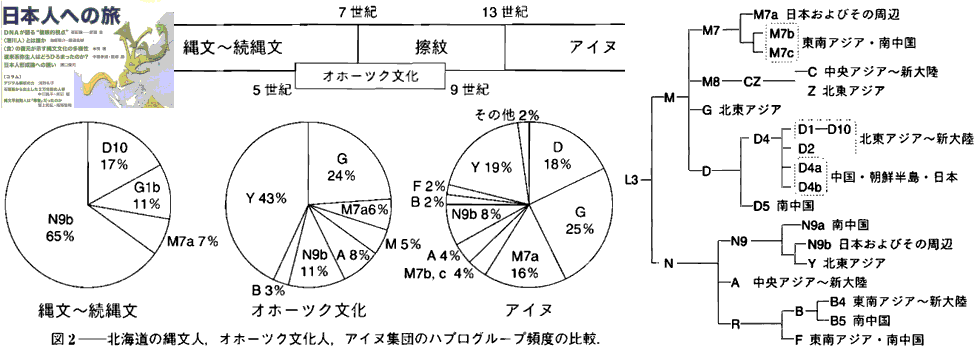
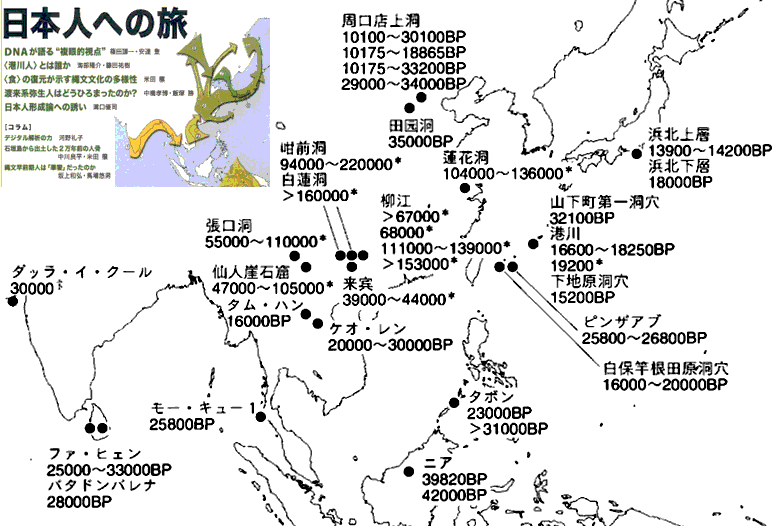
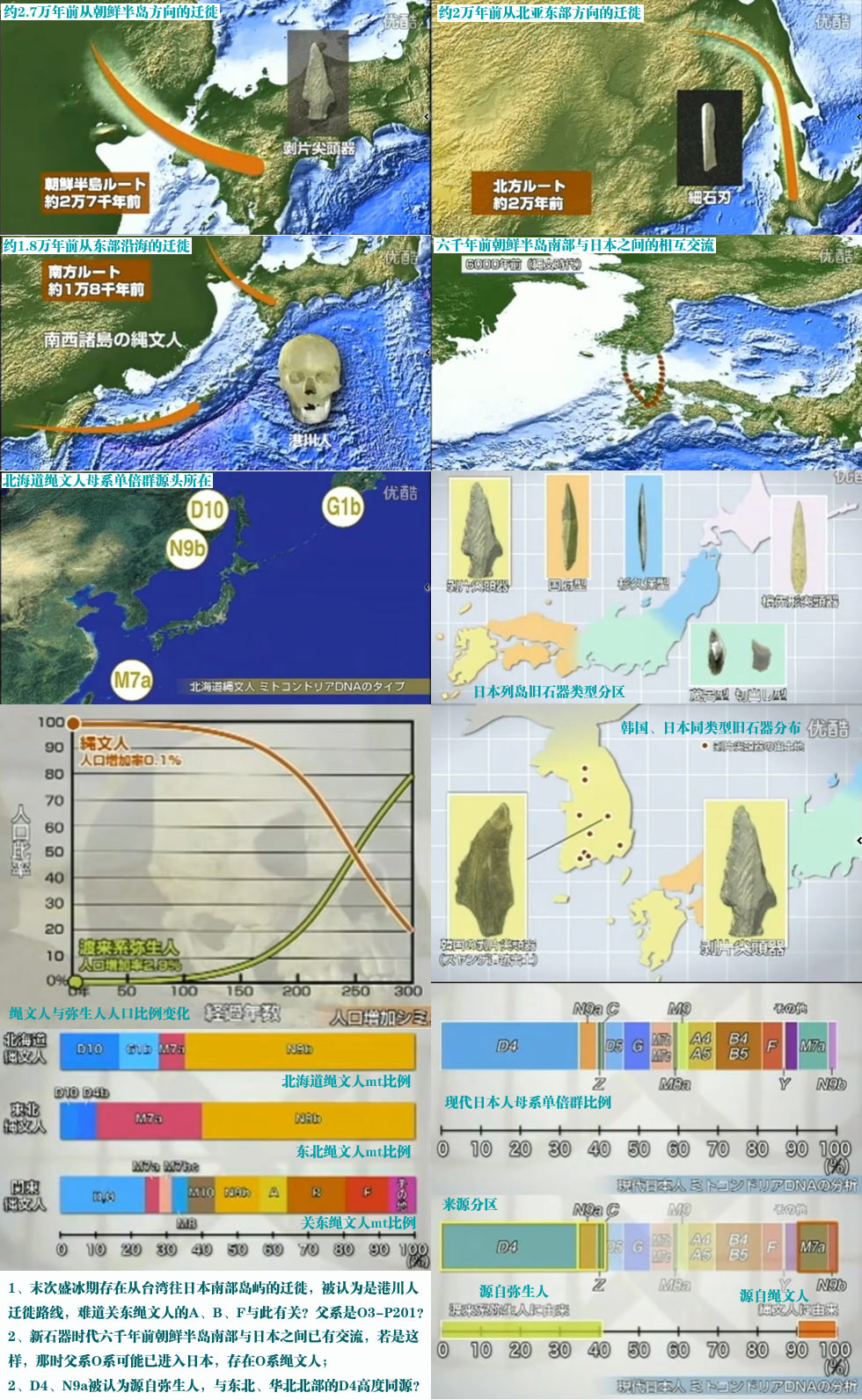
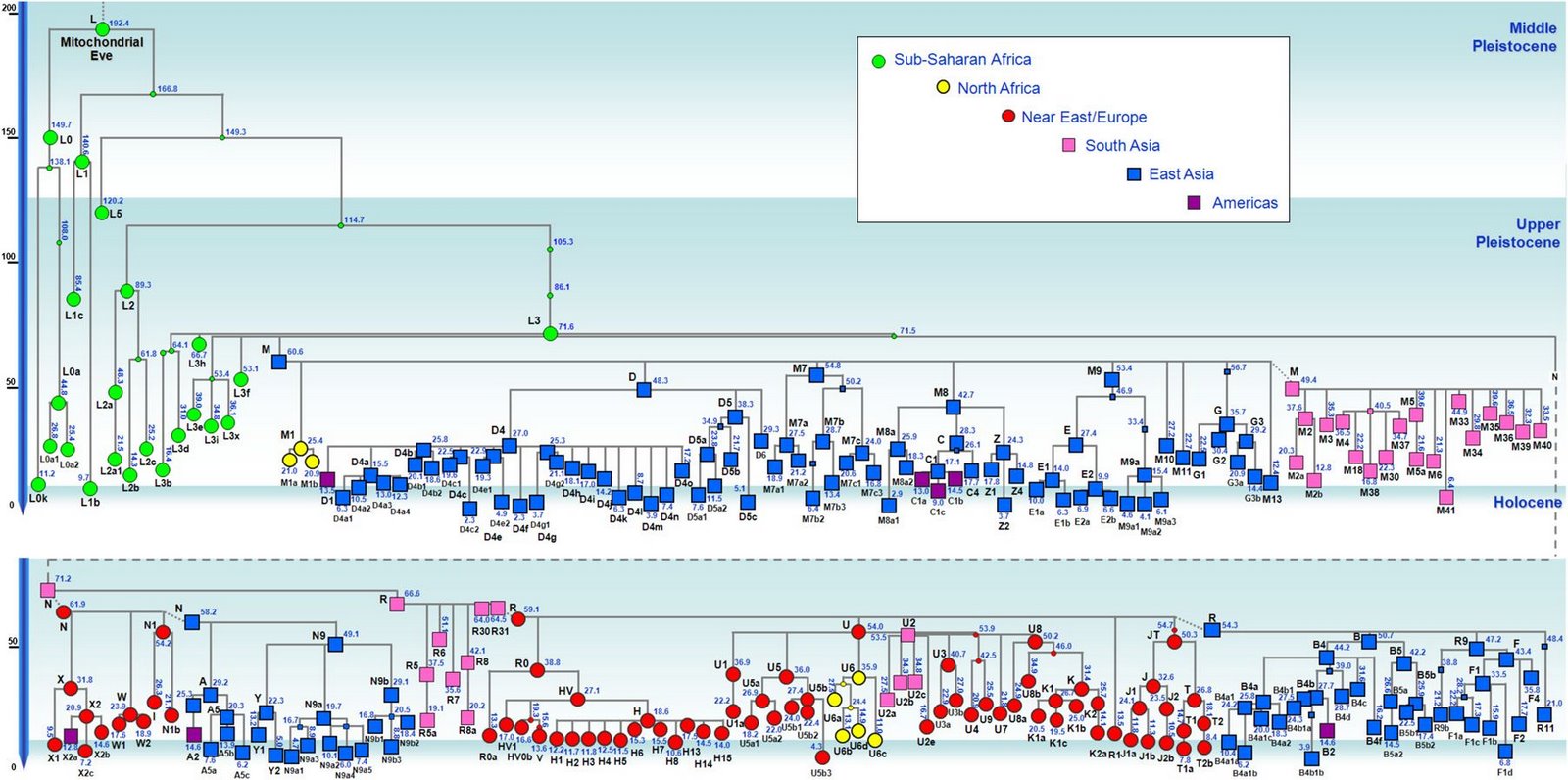
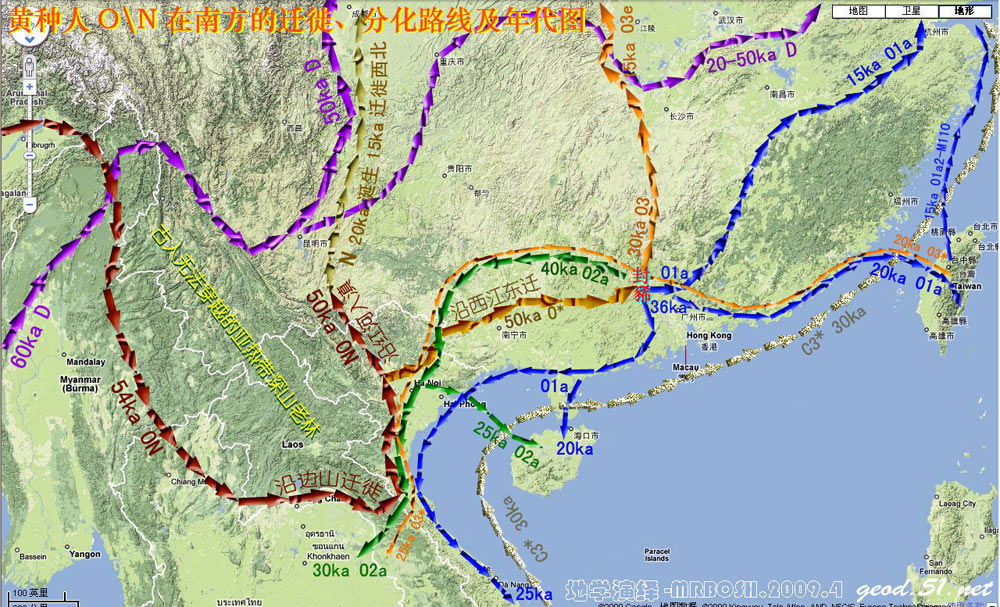
对日本绳文人始祖地的推测 绳文人的父系主要是D2与D*,另有20%左右的C3与C1(C3、C1应从日本北方岛链南下,这里不作讨论);其母系不但有当今东亚北方人群常见的D4、D5、A、G、M8a、M10等,还有南方O系人群常见的B4、B5、M7b、M7c等,仅在日本出现高频的M7a及N9b可能诞生于北方东部,但他们的祖型应是南方起源的,F可以看作是南、北均有的中性成员。这些成员的分歧年代大多不晚于2.5~3.0万年前。根据这些组份及分岐年代推测:绳文人父系D人群的始祖地可能在长江流域。5万多年前,早东亚人父系D人群翻越横断山脉进入云南,然后沿长江而下,D4、D5、G、M8a、M10等应为父系D的原配,而南方O系人群常见的A、B4、B5、M7b、M7c、F有可能来源于云南一带的黄种人父系ON-M214人群,当然,也不排除绳文人母系形成于盛冰期从北方缩回长江流域的父系D人群,然后在湖南一带混入南方人群的A、B4、B5、M7b、M7c、F等。 如果把湖南道县玉蟾岩遗址测得的最早年龄作为O3、也则O系进入岭北的最早年代,O系进入长江流域的年代大约在2万年前,而在此之前,长江中下游还分布着5~2万年前的旧石器晚期遗址,如贵州的马鞍山北洞遗址(20-18ka),湖南的石门人(20ka)、十里岗、乌鸦山、石门燕儿洞遗址,湖北的鸡公山(50-20ka)、伴峡榨洞(20ka)、樟脑洞遗址,江西的新余遗址,安徽的水阳江遗址,江苏的下草湾(50-40ka)遗址等,这些遗址可看作是早东亚人父系D人群在长江流域活动的证据,因为它们当中有末次间冰期遗址,所以,从云南沿长江而下的父系D人群肯定有,但也不排除有一部分是由于到了盛冰期北方气候寒冷,从北方撤回华中地区的人群。当今南方部分瑶族、畲族等的底层就有一定比例的D(父系)。至于为什么瑶族、畲族等中的D是D*与D1,而绳文人的D主要是D2,也有少量D*及D1(见兰海《Y染色体单倍群D在东亚的分布及其意义》.现代人类学通讯.2008)?那是因为当时D2还没诞生,D2-P37.1的估算年代大约在2万年前。 比父系D人群稍晚,ON-M214人群也很早,大约在5万年前就已经进入到云南地区,但父系N大约在2万年前才分化出来,黄种人ON-M214与早东亚人父系D在云南杂居了近3万年前时间,如果从现今父系的N人群与D人群上看,N与D基本算是不曾互混,但北方父系N人群所对应的母系M系比例比N系还高,日本绳文人母系中A、B4、B5、M7b、M7c、F等南方人群成分的比例也不低,从中看出他们之间在母系上是有交流的,这种情况很令人费解,或者是在早期人数还较少的时候,他们在云南发生过互抢对方女人的事件,发生了族群战争,把对方男人杀掉,抢夺了对方的女人做老婆? ON-M214是个很奇怪的单倍型,他作为父系的N与O的祖型,不仅在当今的父系N人群中稀少,更是在O系人群中罕见,也许是因为年代太久远,在历史的长河中绝大部分因漂变而消失所致,也许当初他们曾混入到早东亚人D人群当中,但也因漂变的原因,后来都消失了。 早东亚人父系D人群在旧石器晚期为游猎族群,过着无定居生活,在追逐猎物的过程,迁移速度极快,人口数量不一定很大,但分布范围极广,能在不到1万年的时间里遍及整个东亚北部,大约4万年前就已到达北极圈。他们是如何到达日本列岛的,有两条路线的可能:一是在盛冰期低海平面时,沿着中国东部古海岸直接进入日本,但此路线要经过无石器利用的宽阔无比的泥沙质平原或海岸滩涂,其可行性如何尚无考古方面的依据;二是迁徙东北后经朝鲜半岛进入日本,沿途留居大陆(包括朝鲜半岛)的后来在盛冰期结束后被黄种人消灭,而迁入日本列岛的由于冰后期海平面的上升、与大陆隔绝则生存了下来。 小 结 现代日本人基本是由绳文人与弥生人的延续及混合而成,两者父系的比例大约为2:3,而母系绳文人比例可能更高。 若按迁入时间先后及父系单倍群,大致可分为四批: 第一批是在2.5~1.5万年前末次盛冰期日本列岛与大陆相连时经朝鲜半岛迁入,父系单倍型开始是D*,进入日本后分化出D2,母系单倍型不仅有北方人群常见的D4、D5、A、G、M8a、M10,也有南方人群常见的B4、B5、M7b、M7c、F等,还有在日韩地区才出现高频的M7a,他们的后裔约占绳文人父系的80%。 第二批是在盛冰期结束后、主冰期结束前的大约1.3万年前,当时海平面已上升,朝鲜海峡已不能通行,但俄罗斯远东与日本之间的岛屿基本还是相连的,他们带着细石器技术沿岛链进入日本,早期的制陶技术可能也是从这个方向进入,他们的父系单倍型是C3及C1,母系单倍型是N9b、Y、G等,其中N9b可算是日本特色(俄罗斯远东也有零星分布),而Y是判定其从北方进入的依据(Y的高频区在俄罗斯远东沿海一带),他们的后裔大约占绳文人父系的20%。 第三批在3500~2800年前进入日本,他们是来源中国浙江及山东一带的稻作农业传播者,父系单倍型主要是O3系,也有O1a-M119,他们驾舟沿海岸或经朝鲜半岛进入日本,估计他们的后裔约占现代日本人口的10%左右。 第四批是弥生人,大约是在2300年前从朝鲜半岛南部东渡日本,他们的父系单倍型主要是O2b(O2b1)及O3(O3e),母系单倍群则很复杂,不仅有北方人群常见的D4、D5、A、G、M8a、M9、M10等,也有南方人群常见的B4、B5、N9a、M7b、M7c、F等,他们的祖先可能是分多批次从中国东部到达朝鲜半岛的,既有1万至5千年前的,也有晚于5千年前的。弥生人后裔再加上秦汉以来东渡日本的,约占现代日本人父系的50%。 另外还有在秦汉时期从中国北方东渡日本的,如传说中徐福率三千童男童女为秦始皇出海寻找仙药没有回来,可能是到了日本。北方东渡人群父母系单倍群应与当今北汉差别不大,父系应是O3(M134、M117及002611),而母系主要是D4、D5、A、G、M8a、M9、M10,母系N系的B、F、N9a比例应比现代稍高。秦汉东渡人群占日本人口比例估计不大,因为日本父系O3总体比例也只有23%。
《日本列島人の起源に迫る~私達は何処から来たのか~》视频截图 |

{kind=link}
{kind=link}