![]()
|
|
|
|
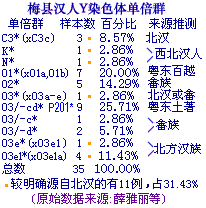
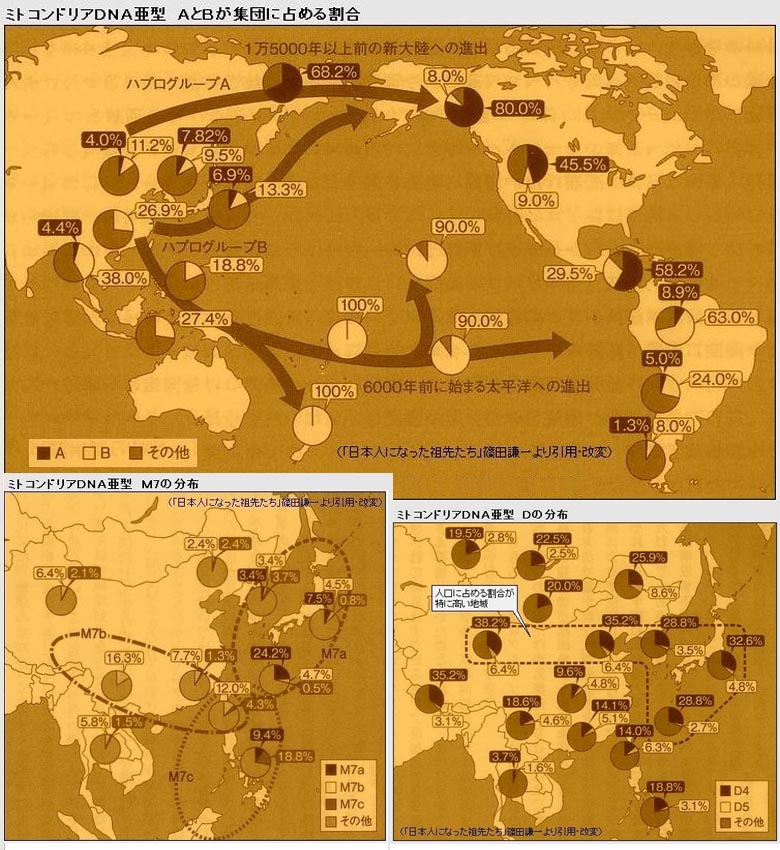
地学演绎 莫如波 2011.12.28 线粒体A单倍群分布概况 从文波博士学位论文的数据中看到,较高比例的A主要分布于北方汉族、北方少数民族及北亚民族当中,如甘肃汉族17.0%(8/47)、武汉汉族16.7%(7/42)、新疆汉族10.6%(5/47)、西宁汉族9.1%(4/44)、西安汉族8.2%(7/85)、朝鲜族14.6%(7/48)、东蒙古12.3%(7/57)、土族11.4%(4/35)、日本人10.1%(9/89)、蒙古族8.3%(4/48)、韩国人7.3%(23/313)、西蒙古6.9%(4/58)等;西南(或西部)少数民族大多亦有较高比例甚至比例更高,如傈僳族33.3%(11/37)、怒族30.0%(9/30)、青海藏族21.4%(12/56)、普米族14.3%(5/35)、哈尼族12.1%(4/33)、云南藏族12.1%(12/99)、彝族11.5%(10/87)、土家族9.3%(9/97)、湖南苗族9.0%(9/100)、纳西族8.9%(4/45)等;南方汉族、岭南少数民族以及部分西南少数民族A比例低或没有,如广州汉族(0/60)、浙江汉族(0/30)、广西汉族4.5%(1/22)、壮族1.7%(2/119)、瑶族2.2%(6/275)、台湾原住民0.5%(1/176)、水族(0/30)、佤族2.3%(2/84)、布朗族(0/28)、拉祜族(0/82)等[2,文波]。从另几份数据中看到,隔绝环境下的台湾原住民672个样本竟无一例A,海南琼海黎族100例也未见有A,没有A的还有广州人(0/68)、湛江人(0/30)、南昌人(0/23),客家祖地福建长汀人(0/54),见>>>、>>>。据陈致勇,A在东南亚分布极为有限,泰国有一定比例,马来人低频,东南亚岛屿未发现有A[1,陈致勇]。
A与父系单倍群的关系 结合以往的分析,母系有A单倍群的父系族群有O3系,如北方汉族;有N系,如长城地带古华北人种戎狄的古DNA数据[3,赵欣];有C3系,如北亚人群古DNA数据[4,周慧],还有北美土著有A2;有D,如藏族、关东绳文人;据陈致勇,中国东部沿海A原始型及A4原始型频率较高,如马桥原住民、福建莆田人、潮阳人、潮州人等[1,陈致勇],马桥原住民的父系主要是百越O1a(P203)[5,李辉等],莆田人、潮汕人的父系主要是中原O3(M134、M117)[6,李辉]。最欠缺A的人群是父系的O2a(M95、M88)人群,其次是岭南原先的O1a(P203)人群,南方的一些O3土著也欠缺A,如孟高棉-苗瑶系统、台湾阿美族等。可以大胆推测,母系A能贯穿北方父系D、N、O3、C3等所有东亚起源的人群,应与末次盛冰期沿长江发生过的东西向交流有关。 A的类型 首先是她的原始型A*,从原始型当中分化有两大族系A4*与A5*,还分出如A-16234、16293C及A8等支系,余下的可归为未定义类型[1,陈致勇]。A4*主要分布于东亚大陆,主要由她的原始型及未定义类型组成,一些大的支系有A4-16274、A4-16124、A4a*、A4b*、A2等;A5*主要分布于中国东部沿海及日本、韩国,A5*未找到早期类型,所见的是她的三个分支A5a、A5b和A5c[1,陈致勇],A5a见于日本及朝鲜半岛,东亚大陆极罕见[1,陈致勇],其应诞生于旧石器晚期的日-韩,A5b见于中国东部沿海,亦见于日本南部[1,陈致勇],可能在大约2万年前诞生于中国东部沿海,盛冰期低海平面时部分迁徙日本,A5c年代只有几千年[1,陈致勇],可能诞生于东北亚沿海,低频地分布于北亚东部及日本。(本段主要为陈致勇《东亚人群线粒体N系单倍群的迁徙分析》中的内容,所谓的原始型是指在高变1区特征上与系统发生树推导出的单倍群祖型完全一致的单倍型,未定义类型是指在该单倍群的高变1区数据中剔除原始型及其主要分支后余下的单倍型) A单倍群的诞生年代 陈致勇采用高变1区数据测算,得A的分化年代为39.6±13.2ka,分布于中、日、韩及北亚的A5b的分化年代为16.5±9.4ka,采用编码区数据测算,得A5a的分化年代为15.4±4.9ka[1,陈致勇];在一张母系支系年代图上,A的年代为29.2ka、A5b为13.9ka、A5a只有7.6ka,Hong-Xing Zheng, Shi Yan等估算的A的年代约为20ka[7]。从A单倍群分布特点分析,岭北的O3系统都有A,但台湾原住民的O3(P201*)人群没有A,半岛孟高棉及瑶族的O3系统欠缺A,原先的O1a系统及O2a系统没有A,可见,A应该诞生于O3系统最早期的一个将向北方迁徙的分支,A诞生的年代应晚于父系O3诞生的年代,则大概不早于3万年前,合适的时段大致为30~25kaB.P,她是在早期所在部落离开O3(P201*)人群以及原始孟高棉-苗瑶的O3系统分出以后才诞生的,诞生地点陈致勇根据A单倍群的分布频率及内部变异频率在各族群的分布情况,锁定在桂东北[1,陈致勇],也则贺江中上游地区,诞生以后就进入湘南;A5a见于日本及韩国,东亚大陆极罕见[1,陈致勇],可算是日韩特征单倍群,应该是末次盛冰期低海平面时A*迁入日本列岛以后诞生的,年代应不晚于1.5万年前,陈致勇所测算的“15.4±4.9ka”应可接受;A5b的分布超出日本,应诞生于A*迁徙日本之前,“16.5±9.4ka”也说得过去。母系支系年代图上的A5b尤其A5a共祖年龄过小,可能与样本所囊括的支系偏少有关。 A原始型与A4原始型的分布应与她们早期的迁徙聚集有关 陈致勇统计:“A原始型分布频率最高的族群是广东潮阳人5.3%,其次是广西东北部的富川瑶族3.2%和罗城仫佬族2.6%,以及湖北汉族2.4%和内蒙古阿荣旗朝鲜族2.1%。A原始型在东亚的分布呈分散的点状,在江苏汉族、呼和浩特蒙古族、广西拉珈人、泰国曼谷人、本土日本人、韩国人中都有发现。”[1,陈致勇]陈致勇分析认为:“在A单倍群原始型发生频率上叠加A单倍群群体内部变异频率得到的叠加分析图。可以看到在中国境内单倍群分布频率和内部变异频率重叠高发的区域有两个:一是广西东北部地区,据统计这里内部变异频率最高的族群是罗城仫佬族7.4%,邻近的广东怀集标人更高达8.8%;二是湖北东部地区,武汉人的内部变异频率是4.8%。在境外,日本南部和泰国南部地区也有重叠的次高发区,日本宫崎县人的内部变异频率是5.6%,泰国曼谷人的变异频率是3.0%,显然都不如广西东北部和广东西部一带的变异频率高。据此我们基本上可将A单倍群的最初分化地点锁定在广西东北部一带。”[1,陈致勇]陈致勇认为:“A4原始型分布范围很广,并且呈现相当离散的状态,表明早期分化历史已很久远。分布最密集的地区是在中国东南沿海和日本南部一带,这可能就是A4第一波扩张所及范围。其他分布较多的地区在图中呈现为灰色,这些可能是伴随A4其他分支迁徙的原始型所留下的遗传印迹。据本次研究的结果,A4原始型最高发的是广东潮州人,发生频率为8.3%。其次是西双版纳彝族6.3%、福建莆田人6.1%、贵州赫章县彝族5.0%、和日本南部宫崎县人4.0%。A4原始型也见于韩国人中(2.3%),但在日本中部人中没有找到。值得注意的是,A4原始型出现在百越起源的上海马桥原住民中(5.9%),在51个阿伊努样本中也发现1例A4原始型。这暗示A4早期扩张主要走沿海路线而且年代十分久远。”[1,陈致勇](注:潮阳人、潮州人[8]、福建莆田人为中原移民,数据所代表的应是古代中原河南的情况{8])
日本关东绳文人的父系应是D(D2) 绳文人是指万余年前至3000年前的日本古代人,关东绳文人的母系有较大比例的A、B、F,也有一定比例的M7b、M7c[9],当中B、F、M7b、M7c是中国南方人群最常见的单倍群,这很容易使人想到,关东绳文人可能是新石器中期移民,对应的父系应是O系,是O2b或O3。从日本人的数据中看到,北线北海道绳文人母系单倍群为N9b、G1b、D10及M7a[10],当中M7a应为来自日本南部的移民,北线应是没有A、B、F、M7b、M7c的,可以推测,A很可能是与B、F、M7b、M7c一块来自中国东部沿海,只要把A的源头及年代确定了,关东绳文人的源头也就可确定。A5a见于现代日本人及朝鲜半岛人群,东亚大陆罕见,与M7a一样可算是日本及韩国的特有单倍群,A5a的共祖年龄为15.4±4.9ka[1,陈致勇],鉴于日本南部宫崎县人A原始型内部变异频率较高(5.6%)[1,陈致勇],A4原始型也在日本及韩国南部有较高频率[1,陈致勇],认为A*可能是先进入日本南部,诞生了A5a以后在海平面上升前再流向朝鲜半岛,因此,A在末次盛冰期低海平面时应已进入日本,关东绳文人的A及B、F、M7b、M7c很可能是旧石器晚期遗民。她们所对应的父系是什么呢?占现代日本人父系最大份额的是O2b与D2,再者是O3,剩下的其他类型比例不高。O2b万余年前大概诞生于北部湾一带,其未能在盛冰期低海平面时进入台湾成为台湾原住民的一员,因此,他们也不太可能在末次盛冰期迁入日本列岛,目前多数认为,O2b大约在2300年前才从朝鲜半岛南端东渡日本,是弥生人的主体成员;至于O3,以前曾有分析,在O3的几种常见单倍群当中,O3a3c-M134*、O3a3c1-M117及O3a4-002611早期主要聚居于广西与湖南,他们向中南半岛、中国北方及东部沿海地区的迁徙扩张都是盛冰期结束以后的事,年代不早于1.5万年前,他们虽在日本人当中有一定比例[11,Nonaka],显然 母系A诞生于贺江中上游的父系O3族群,她们又是如何很早就随父系D人群迁徙日本?如何很早就随C3*迁徙北亚东部并到达北美?由于A不是诞生于华南沿海或东南亚沿海,与父系C3*在南方没有交汇点,A也不是诞生于西南云南一带,初始与父系D也没有交汇点,迁徙途中的“碰合”似乎是其唯一的融入方式。从A的原始型及A4的原始型分布推测,他们在贺江中上游诞生后,很快就进入了湖南,然后顺长江而下迁徙到东部沿海地区,当今东部沿海A原始型及A4原始型频率较高是存在这种迁徙的反映。迁徙过程,她们一部分碰到了更早就生活在长江流域的父系D游猎采集族群,被劫抢成为父系D的女人(最早进入湘南时可能就遭到了食人族的伏击,男人O3被吃掉而没有留于D族群之中),后来他们部分迁徙日本成为关东绳文人的祖先;又有一部分在长江下游沿海一带,碰到了从华南沿海向北迁徙的父系C3*人群,她们伴随C3*迁徙至东北亚,并有一部分(A2)在1.4万年前跨越白令海峡到了北美。从另方面,我们可以从原居长江中游的荆蛮的后裔及相关人群中找到早期父系D的踪迹,部分人群混有较高比例的D,如广西金秀拉珈瑶52.2%(12/23)、金秀茶山瑶50.0%(5/10)、广西隆林徕人37.9%(11/29)、金秀瑶族30.0%(3/10)、云南宁蒗彝族18.4%(7/38)、江西兴国畲族18.2%(4/22)、浙江景宁畲族18.1%(13/72)、广西富川平话汉族17.9%(5/28)、富川平地瑶16.1%(5/31)等(数据来自兰海)[14];再可从长江下游或原居长江下游的人群中找到早期父系C3*的印迹,部分人群C3*比例较高,如上海汉族22.22%(4/18)、江苏汉族15.38%(6/39)、安徽汉族13.46%(7/52)、湖北土家族30.77%(8/26)、贵州侗族29.63%(8/27)、湖南侗族22.22%(10/45)、贵州土家族21.21%(7/33)、广西仫佬族18.18%(2/11)、贵州水族13.79%(4/29)等(数据来自Hua Zhong, Hong Shi等[15])。 西南少数民族的A大部分源自长江中游地区,年代较晚,属新石器时代移民 如集中分布于云贵地区藏彝系民族中的A-16234,16293C,其在云南西双版纳彝族和贵州赫章县彝族最高发,比例分别为12.5%和10%,在云南藏族、白族、普米族、纳西族以及西藏藏族中发生频率为2.2%~4.7%,A-16234,16293C的共祖年龄为7.2±3.8ka,从其分布特点不难看出,它应该是先进入彝族母系遗传中,而后再随着彝族的迁徙进入到周边族群;又如A4-16274,其为A4在中国境内最大的支系,主要集中在云南藏彝系民族中,其中频率最高的是怒族30.0%、傈僳族16.2%,其次是普米族8.3%和撒尼人(彝族支系)6.5%,在湘西过山瑶中的频率是4.2%,在云南、青海藏族中的比例为3.6%~4.0%,在华中、华东汉族中低频,发生频率从1.3%至2.4%不等,A4-16274的共祖年龄为3.6±1.6ka;再如A4-16124,其在西部藏彝系民族中较常见,与A4-16274的分布特点相似,只是发生频率相对较低,最高发的是云南傈僳族13.2%,次之贵州赫章县彝族5.0%、湖北汉族4.0%、云南撒尼人3.2%、云南藏族2.9%和西藏藏族2.9%,A4-16124的共祖年龄为3.4±1.7ka,大部分样本属于原始型,暗示其在华中地区形成,向西南地区迁徙发生在晚近期;A4未定义类型主要集中在内陆特别是华中地区,最高频率湖北恩施侗族高达30%,依次是甘肃汉族15.6%、贵州沿河县土家族10.3%、贵州赫章县彝族10.0%、青海藏族8.9%,从分布图看,其应是先在华中地区(准确地说是在湘鄂渝交界区域)扩散,再向西部藏彝走廊和西北地区扩张,很可能是较晚期的一次大规模扩散所留下的痕迹。[1,陈致勇](本段摘编自陈致勇《东亚人群线粒体N系单倍群的迁徙分析》中的内容)
主要参考文献 [1].陈致勇.东亚人群线粒体N系单倍群的迁徙分析.现代人类学通迅.第三卷.2009年; ①.mt-A诞生于3~2.5万年前的贺江中上游地区,孟高棉-苗瑶族群欠缺A,由此推测,原始孟高棉-苗瑶的O3系统(M117+M7+M134+M122*)在母系A诞生前已向西分出。 ②.伏羲族系大约诞生于1.1万年前,北方华夏族群、东夷族群及兴隆洼支系当属伏羲族系,当中母系有A,湘西北彭头山文化与贾湖文化有同源关系,因此,彭头山人群也应属伏羲族系,其应是一群以O3(M117+M134+002611)占绝对优势、混有少量O2*-P31、母系有A的族群。 ③.原始孟-苗族群不属伏羲族系,新石器早期的彭头山人群应还没有孟-苗系统的M95+M88+M7。彭头山遗址见二次葬,还未出现屈肢葬,新石器早、中期广西骆越流行的屈肢葬见于湘西南七八千年前的高庙遗址及鄂西六千多年前的大溪文化。大溪文化的古DNA以O3-M7为主,当中有O2a。由此判断,七八千年前,原居广西北部的原始孟-苗开始进入湖南并向北过长江到达鄂西,过程孟-苗父系的O3-M7、O2a(M95、M88)融入到湘西鄂西伏羲族系当中并形成荆蛮族群。 ④.高庙文化很发达,陶器上的八角星刻符可能是伏羲八卦的某种形式,“高庙文化最初(约7800年前)出现的八角星纹以及獠牙兽面、鸟载太阳等图像,在较晚时期逐步流行于长江中、下游地区及黄河流域,甚至更远的地区。在洞庭湖区的坟山堡、汤家岗遗址,太湖流域的崧泽文化,安徽含山凌家滩遗址,山东的大汶口文化,以及辽宁小河沿遗址南台地,出土的某些陶器或玉器上均见到与高庙文化完全类同的八角星纹。”后来的城头山遗址、凌家滩遗址等都发现有类似于高庙遗址的、与墓地相结合的祭坛(见《高庙新石器时代遗址》)。贵州远口镇坡脚遗址、广西桂林甑皮岩遗址和平乐纱帽山遗址等均出土有其主体特征属于高庙文化的遗存。之后,来自洞庭湖区的大溪文化对更大区域产生了强势覆盖(见贺刚,陈利文《高庙文化及其对外传播与影响》2007)。荆蛮族群可能是在伏羲族系、伏羲文化的基础上融入了大量孟-苗人群所形成的,主要爆发于有M95、M88、M7的高庙人群后裔大溪文化期,新石器中晚期荆蛮族群遍及长江中游地区。 中国社会科学院民族学与人类学研究所吴安其《长江黄河流域和华南地区的古代语言》(现代人类学通讯.2012年.第六卷)文摘:“汉藏、南岛和南亚语系有着一系列形式相同的形态特征,如*s-使动前缀,自动词*m-前缀,*-n名词后缀等,以及一些基础词有相同或相近的词根。它们原本可能来自古华南语(或称华澳语)。汉藏、南岛和南亚诸语是在早期华南语言的基础上形成的。”“八九千年前汉藏语北上淮河和黄河流域形成早期的汉藏语”“古汉语的词跟藏缅语、南岛语的关系最为密切,复辅音声母对应藏缅语的复辅音声母和南岛语的复音词头两个音节的辅音。”“湘西地区原本是南亚文化,后为苗瑶人所居住,这与苗瑶语中包含着南亚语底层的情况符合。(颜其香,周植志.1995)”“湘西原有的土著文化战国以后由湘西北开始渐为湘北文化,即古苗瑶文化所取代,现代苗瑶语不同支系中保留的南亚语的成分,这是原湘西等地南亚语的遗存。” |
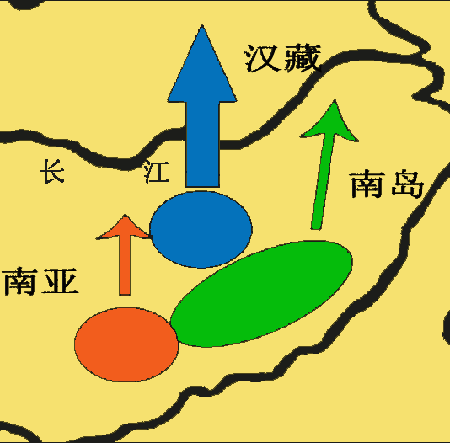
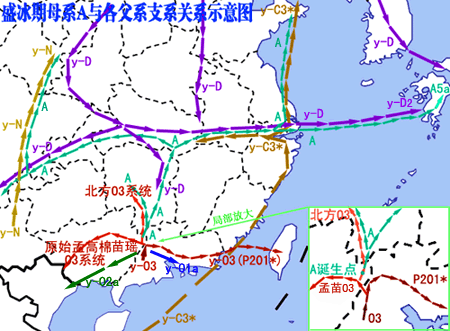 进入盛冰期,由于气候寒冷,原居北方的父系D游猎族群回流南方,而居岭南的O3人群由于人口的增加从桂北进入长江流域(湖南),云南父系N也已诞生并开始扩散,C3*沿海岸从南往北迁徙。结果多种父系单倍群的人群沿长江一线发生了会合与交流,也正是这种横向的、东西向的交流,导致了母系A单倍群融入到长江沿岸父系分别为D、N、O3、C3等的各路人群当中,再随着冰后期的北迁扩散到整个东亚北部及更多地区。
进入盛冰期,由于气候寒冷,原居北方的父系D游猎族群回流南方,而居岭南的O3人群由于人口的增加从桂北进入长江流域(湖南),云南父系N也已诞生并开始扩散,C3*沿海岸从南往北迁徙。结果多种父系单倍群的人群沿长江一线发生了会合与交流,也正是这种横向的、东西向的交流,导致了母系A单倍群融入到长江沿岸父系分别为D、N、O3、C3等的各路人群当中,再随着冰后期的北迁扩散到整个东亚北部及更多地区。")
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}