![]()
|
|
|
|
发于论坛上的内容再整理 关于亚洲东南几路最原始人群的分化 地学演绎 莫如波 2013.7.21
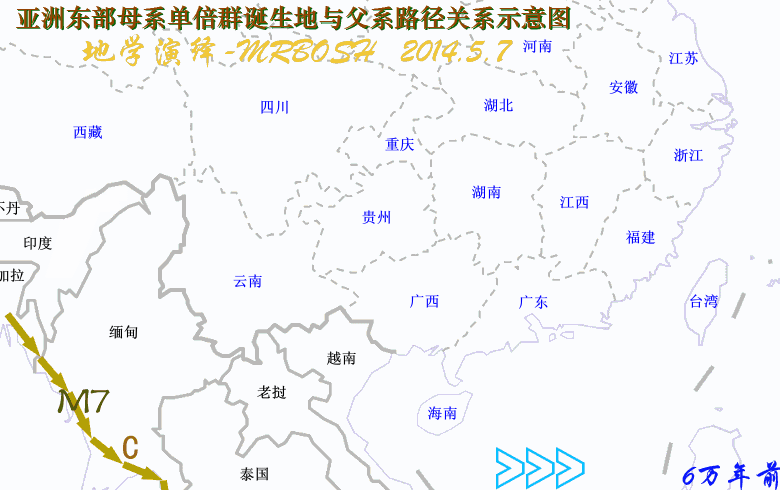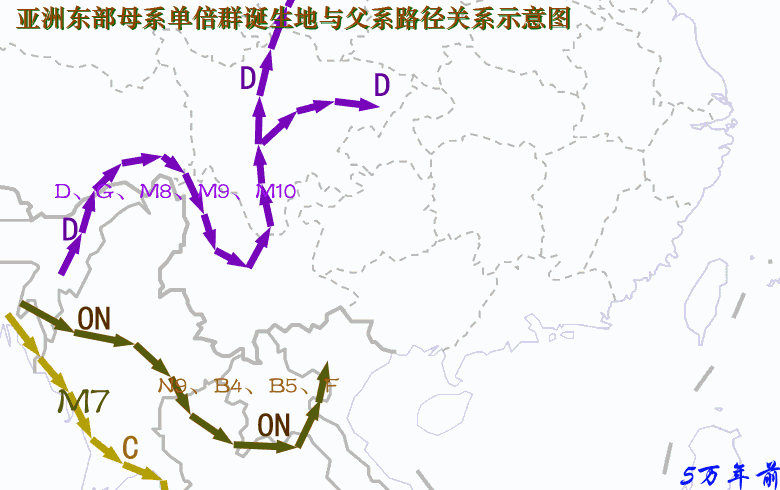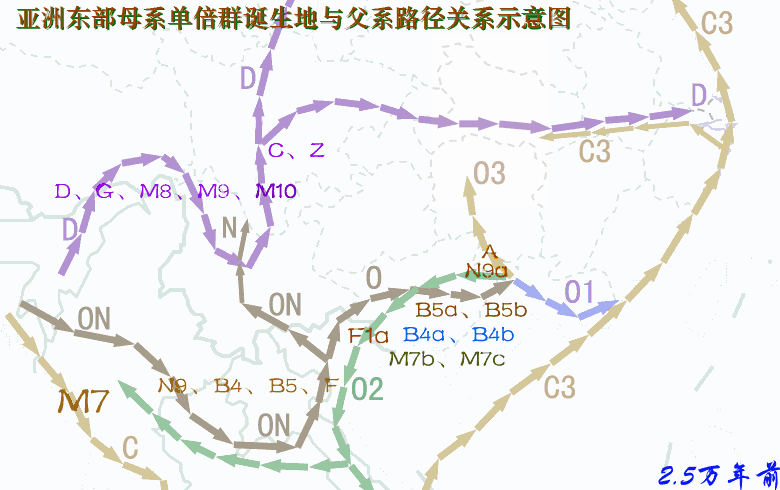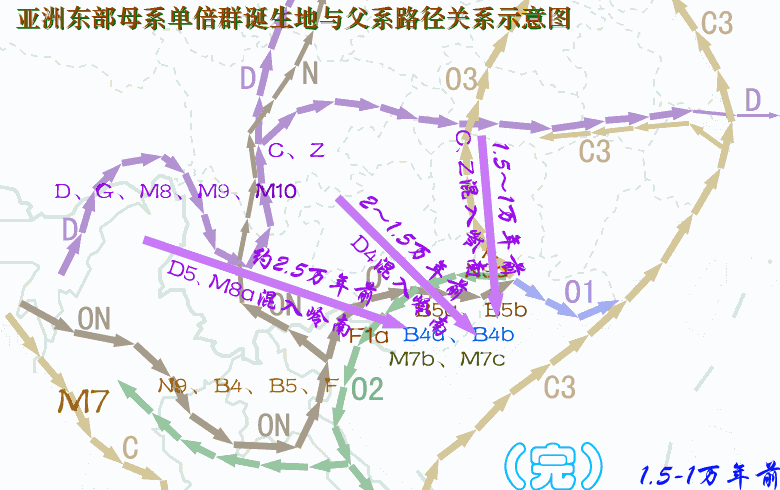
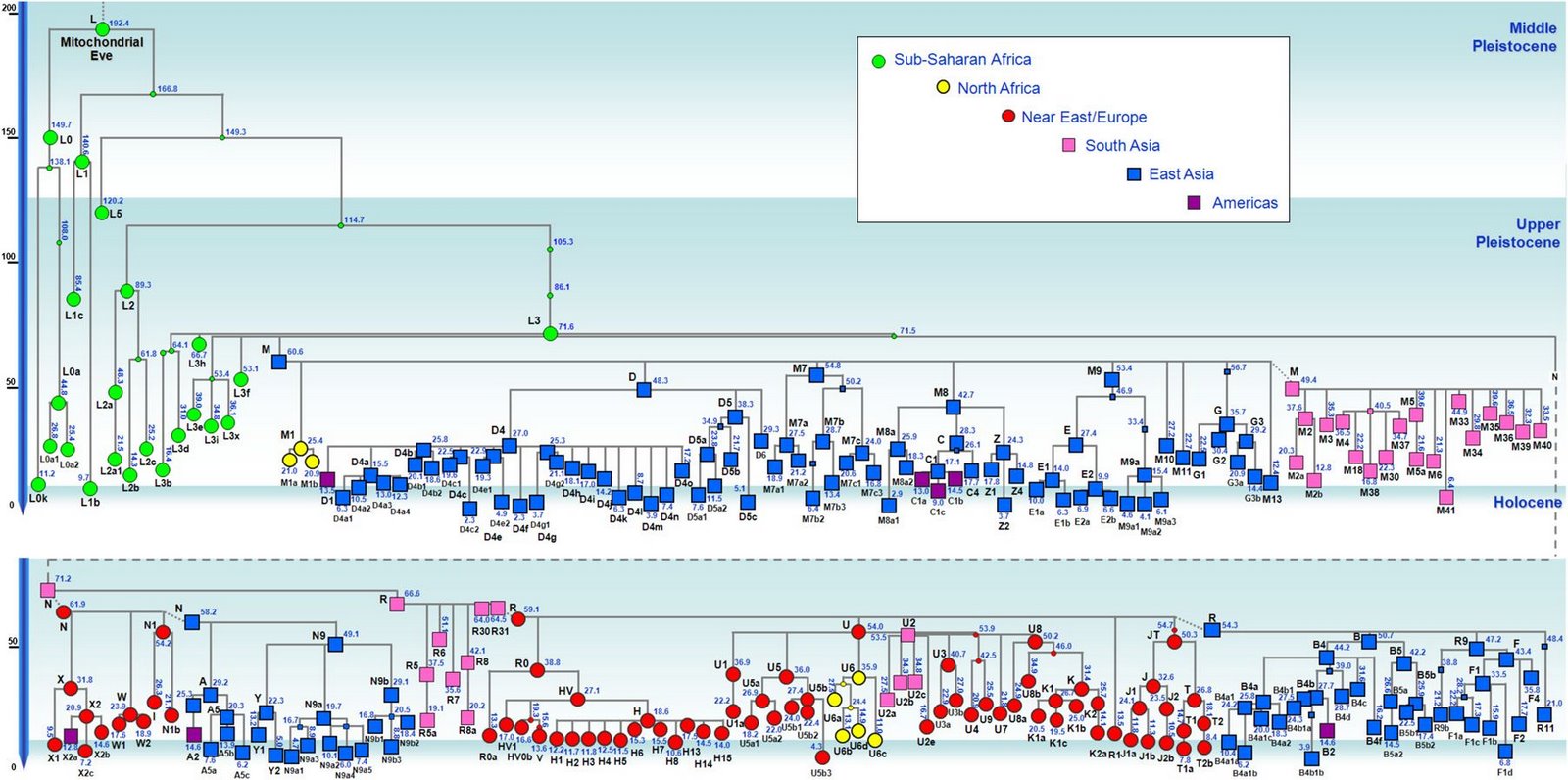
现代人祖先从非洲迁徙亚洲东南部,最早期按时间先后可分为三大支系:一是约六万年前东南亚沿海支C,其中C*、C3初期居中南半岛沿海,C3对应的母系由于后来与O系相混,已难以分辨;二是五万多年前西线内陆支D,从印度东北拐进藏东及云南,三五万年前这一支大概又分为两个分支:一个分支沿西北黄河上游方向迁徙,母系单倍群有D、G、M8、M9、M10等,另一分支迁徙长江流域,有一部分再迁日本,并形成关东绳文人,母系除原配D、G、M8、M9、M10等外,还混进了A、B、F等N系成员以及M系的M7b、M7c,混进部分很可能来自O系;三是五万年前从北部湾迁徙岭南的O系,四-三万年前先后分化出O2、O1、O3,母系是N系的B、F、N9、R9等以及M系的M7b、M7c,后来在桂北、粤西北、湘南一带又分化出A。另外还有一个次分支NO下游的N,很可能是从北部湾进入云南,初期对应的母系应与O系相同,或F*相对高频,但由于NO在云南在y-D族群的夹缝中混了三万年,后来在N诞生的时候母系已被替换成M系的C、D、G、M8、M9、M10等,这些类型在华北北部长城地带青铜时代的古DNA中可见。 对baiyueren收集数据绘制的母系单倍群频率分布图的看法:尽管不清楚各种单倍群的诞生地在哪里?最早期扩张在什么地方?仅从现代人群分布频率中看,A、N9a可能与湘地或粤、桂、湘交界地区的O3有关,A的年代为2.5~3.0万年,N9a年代约2万年,A在岭南侗台人群及台湾、海南、南岛原住民中少见或罕有,但N9a在这些人群有一定比例,似乎N9a的诞生地或初始扩张地更偏南,或在两广北部。F1a在岭南及南岛都有较高比例,早期对应的父系应是O2a-M95。B4a、B4b对应的父系是岭南百越的O2a与O1a,其最先在广西的O2a人群中扩张,然后又进入东部的O1a人群。B5a、B5b在粤桂湘交界地区及江西一带高频,说明曾在这些地区爆发,但初期扩张地在哪里?从南岛人群亦有较高比例推测,早期扩张应在北部湾广西一带。 A、N9a、B4a、B4b、B5a、B5b、F1a的年龄都在2.5万年左右,很可能都诞生于两广,A靠近湖南,N9a稍偏南大概在桂北一带,两者后来主要随O3进入湖南,然后再向北方迁徙;F1a在广西诞生后,最早一部分在盛冰期随O2a经中南半岛迁徙南岛,更大一部分留居岭南,后来又有一部分随O3入湖南,再迁西南及北方;B4a、B4b、B5a、B5b情况类似,当中B4a、B4b主要构成岭南百越的母系,而B5a、B5b一部分迁南岛,一部分在长江中游发生了大规模的扩张,在荆蛮人群中有较高比例,另外B5b在中原东部有一个相对高频区。至于N9a、B4a、B4b、B5a、B5b、F1a的上游,N9、B4、B5、F的年龄有四万多年(4~5万年),他们的诞生地很可能在北部湾一带。 A诞生后就向北迁徙,岭南侗台人群A低频,海岛原住民未见A,长江流域有大把的A。D4在南方侗台人群中普遍,在台湾及海南原住民中亦有,但南岛原住民没有,所以,判断从西南进入岭南应不晚于1.5万年前,但估计不早于2.5万年前;D5在侗台人群中有,在台湾、海南及南岛原住民中均有,所以从西南进入岭南较早,估计不晚于2.5万年前。M8a的年代是2.6万年,比D5的3.8万年晚了不少,但M8a的分布极像D5,只不过比例小了很多,M8a很可能是伴随着D5一起扩散的,可判断,D5、M8a在南方大规模扩散大约在2.5万年前。G及M9a在侗台人群及台湾、海南及南岛原住民中均罕有,在岭南谈不上扩散;M10在南方少见,主要分布北方河南、河北以及日、韩等地。 北方O3人群母系中比例偏高的M系应与更早居西北及华北北部的采集狩猎族群融入有关。一群父系为N的族群、掌握着细石器技术的采猎族群,北方O3人群偏高的M系大部分来源于新石器晚期至青铜时代的y-N族群,也有一部分可能在新石器早中期,在关中地区已经融入到O3人群当中。 关于O系与C、D的新老关系,D及母系D、G、M8、M9、M*进入西南最早大概在5.5万年前,而ON大概也在5万年前进入西南,O及母系N系在5万年前进入岭南。按“最早”论,所谓的新、老也只差5000年左右。至于C*、C3早期可能居于东南亚沿海,进入东亚大概在3万年前,并很快就沿古海岸迁徙北方及长江下游地区。年代差不了很多,这个老与新似乎意义不大。
最近,国外学者在《欧洲人类遗传学杂志》上发表文章,通过对全球4413名男性当中的K2-M526单倍群的进行分析统计,在东南亚岛屿人群中检测到欧洲支系P1-P27(Q、R)的诸多兄弟分支(相关支系与频率>>>),从而估计,K2-M526初始在东南亚、南岛(南洋)有一个快速分化过程,随后Q、R的祖先P295向西、向北折回中东,迁徙西亚及欧洲等地,如natsuya网友所转载的外国论坛上的示意图(下图),图中,NO-M214亦诞生于中南半岛南部或南洋,然后沿中南半岛东部沿海北迁,进入东亚。Ryan网友在马来西亚人群样本中找到一例NO*,并认为 4.5万年前的西伯利亚Ust'Ishim古NDA pre-NO在谱系树上与NO的距离是比较远的,可能在Ust'Ishim分化出去500~700年之后,NO才在谱系上与目前分布于印度和马来西亚的pre-NO发生分化,这种谱系关系更倾向于NO与其他支系一起经由南亚和东南亚路径扩散。氐羌人后裔网友:“重新看了一下karafet 2015那篇论文的数据,看来P从东南亚起源是跑不掉了。无论是其上游支系,兄弟支系,还是其下游稀有支系P*,都在东南亚。P*在东南亚的分布是:菲律宾Aeta人(在25个样本占28%),Sulawesi(在177个样本占0.6%),Sumba(在649个样本占3.2%),Timor(在509个样本占10.8%),尤其是在Timor中比例更高。从这个角度,P的东南亚起源看来是没什么疑问了。” 大约6万年前,K2与C、D一样,可能都是沿着海岸快速从北非、中东向东迁徙。C年代更早,在海岸势力强大,他们一直沿着海岸迁徙,很快到达澳大利亚及北亚沿海,并跨越白令海峡到达美洲;D首先在印度东北迁向内陆,迁入藏东及云南,最近在菲律宾南部小岛找到了D最早期的并列于D1、D2、D3共同祖先的兄弟分支(据兰海),说明最早在南亚印度沿海,有少量D*沿海岸迁徙南洋;NO-M214可能是第二个适应内陆生活的族群,估计在中南半岛南部已逐渐转向内陆,不然他们也会像C3一样直奔东亚北方沿海,K2a(NO)-M214不一定诞生于南洋,似乎更可能诞生于中南半岛南部,其应是从北部湾进入中国南方的;P1(Q、R)的祖先P295可能是在中南半岛南部或南岛诞生后沿半岛西海岸快速折回到中东、西亚及欧洲的,不然,如果进入内陆,必然在中南半岛发生扩散。(2014.6.8)
马来半岛和婆罗洲北部的原住民9个父系样本中发现N、O的早期分支。马来半岛原住民5个父系样本为:N*—M231/Page91(+),CTS11499/l735/M2291(-)、N*—M231/Page91(+),CTS11499/l735/M2292(-)、I2a2a1b1—L701(+)及2个O2a2a—M188(+);婆罗洲北部原住民4个样本为2个O1b1a1a*—M95(+),F1803/M1348(-)及2个O*—P186/P191/P196(+),M1354(-),M122(-)。当中N*、O*为早期分支,O*见于婆罗洲原住民,是O1a-M119、O1b-M268、O2-M122分化前的早期分支,N*见于马来半岛原住民,是下游大族分化前的早期分支。M188年代较大,突变最大值达2.6万年。但M95+F1803-的分化年代不大,最大值约为1.5万年。因为N*、O*在东亚人群中少见,O*在老挝人群占12%(但也不能排除老挝的O*是P31*),推测婆罗原住民的O*来自中南半岛。M95(+)F1803(-)、M188(+)常见于东亚人及中南半岛人群,由于南岛的O1b系统是青一色的M95(+),M88(-),也没有P31*,判断南岛的M95是在M88诞生前与大陆及中南半岛的O1b系统分开,再混到婆罗原住民当中。还有个I2,应该是近代混入的。这几个检测结果表明,O、N诞生的早期,曾生活于中南半岛,O、N或者诞生于北部湾一带,一部分迁岭南及西南,一部分留居中南半岛及迁南岛;M95是西印尼的主体单倍群,M95是什么时候迁南岛的?按M95的突变年代,似乎不会太早,不会早于1.5万年前,应在中石器时代以来,但有多篇文章似乎在判定M95是在盛冰期低海平面时迁徙海岛的,YFull YTree中M95的年代有没可能算小了?(2018.5.5) 
至此,几路原始人群的主业(主要经济方式)已经跃然而出:D为山区狩猎采摘族群(捕捉动物、采摘野果)、O为河溪渔捞采集族群(捉鱼,摸螺、蚌,采集野果、豆类、野菜,挖掘块茎薯类)、C及其他为海边渔猎拾贝族群(捕鱼、拾贝类)。河滩是古人迁徙的快速通道,海滩是高速通道。O系人群后来从采集活动发展出种植业、农耕业,禽畜驯养很可能也由O系人群发明创造;D族群的狩猎业传给N系人群(细石器文化传播,N是D的女婿),N系人群后来发展出畜牧业。广西已发现贝丘遗址79处,其中洞穴贝丘遗址45处、河旁台地贝丘遗址32处、海滨贝丘遗址3处,洞穴贝丘遗址当中有20处未见陶器,可能年代较早。柳州思多岩、陈家岩、白莲洞1期、鲤鱼嘴1期、来宾盖头洞、崇左矮洞、桂林大岩1期洞穴贝丘遗址认为属旧石器晚期遗址。其中柳州白莲洞C14年龄西10层37000±2000年、西6层28000±2000年、西4层26680±625年,年代距今2.5万年以上;柳州鲤鱼嘴洞穴贝丘遗址可分四个文化期,其中第一期文化期螺壳C14年龄2.1万~1.8万年(据黄启善)。这些旧石器时代晚期洞穴贝丘遗址,说明O系人群早期已经热衷于摸螺捉鱼,到了中石器时代、新石器时代,贝丘遗址就更多,主要分布于中国东部,尤其两广。而D及N在北方,由于气候干冷少雨,河溪不发育,多间歇性河涌,或山势陡峭,河床坡降大水流急,注定不能靠捕鱼及摸螺来维持生计,只能到山上捕猎。
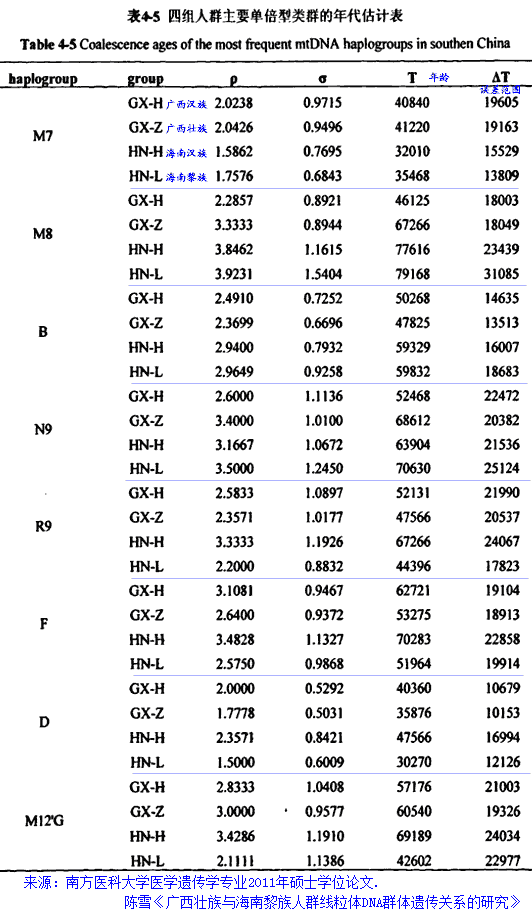
中山大学人类学系乔晓勤、张镇洪等在《华南史前考古若干问题的思考》(纪念黄岩洞遗址发现三十周年论文集.1991)一文中认为,在旧石器晚期-新石器时代初,存在一个经济方式转变的过程,从以狩猎采集为主转变为采集渔捞为主。“在白莲洞遗址,距今2万年以后的地层中螺壳的堆积逐步增多,并有越晚越多的趋势。阳春独石仔、封开黄岩洞、百色的早期全新世洞穴遗址中也有较丰富的螺壳堆积。从世界史前史的材料看,伴随着气候的波动,人类开始减少对大型动物的依赖,转而通过渔猎使水生资源在其食物构成中占有越来越大的比例。与此同时,人类采集活动所涉及的植物种类也越来越多,最终在人口压力等因素的刺激下,开始了植物驯化的过程。从现有的考古材料看,华南山地史前居民在距今3万年至七、八千年这段时期经历了由狩猎-采集经济向食物生产型经济的转变。索尔海姆曾将东南亚的史前文化划分为下列几个主要阶段。①石器阶段(Lithic):这一阶段始于相当于欧洲旧石器时代的早、中期,终于距今4万年前,其主要特点仅见完全打制、技术及类型简单的石器。②竹木器时代(Lignic)(距今四万年-距今两万年)这一时期竹木工具可能是东南亚史前人类工具的主要部分,而石器工业发展缓慢。③结晶化时期(Crystallitic)(距今两万年-公元前八千年)这一时期是东南亚具有独特特征的文化定形的阶段,这个阶段出现了磨制石器与早期种植业。④扩展时期(Extenslonstie)(公元前八千年~公元前后)这一阶段开始于人类由山地向平原,沿海扩展,终于发达的稻作农业及金属的广泛使用(Solheim,1972)。晚更新世的单向打制砾石石器以及与之相应的狩猎-采集经济是本地区文化发展的第一个阶段。冰期过后的全新世初,伴随着贝壳、鱼类,小型哺乳动物成为本地人类的重要食物资源,在以打制砾石石器为主的工具组合中出现了一些小型燧石石器、石箭头与穿孔技术,这是本地区文化的过渡阶段,我们暂称之为“中石器时代”阶段。磨刀石器、低温绳纹陶的出现代表着本地区史前文化第三阶段的开始。在这一阶段上人类已经开始种植包括豆类、块茎类和某些瓜果类的多种植物,这一时期的经济是种植业、渔猎、采集、捕捞共存的复合经济。”-从以狩猎采集为主转变为采集渔捞为主,这种转变究竟发生于何时?是技术上的转变还是整个的换了人(从早期智人换成了晚期智人或从D换成了O)?从母系单倍群的分化上看,大约在2.5万年前应有过一个快速的分化扩张过程,其是否与这种经济转型有关?早于2.5万年前是技术上以狩猎采集为主导还是住着另种狩猎采摘族群?2.5~5万年前遗迹太少,不能排除他们本来就热衷于摸螺与捉鱼。(2014.6.12) 从广西壮族及海南黎族线粒体所追溯到的起源时间来看,他们的祖先四五万年来在岭南的演化应是延续的,2.5万年前应不存在“换人”的假设。 南方医科大学医学遗传学专业2011年硕士学位论文.陈雪《广西壮族与海南黎族人群线粒体DNA群体遗传关系的研究》中的部分结论:广西的壮、汉人群与海南的黎、汉人群在单倍型类群的分布上显示出了极大的相似性,尽管有的类群位于一些分支上,壮族与黎族人群还是共同享有这些单倍型类群,母系基因库都表现出了中国南方人群遗传特征,广西壮族人群与海南黎族人群很有可能起源于共同的祖先,海南黎族很可能是由广西壮族繁衍而来。M8是一个比较古老的单倍型类群,B、R9、N9、F以及M12'G单倍型类群相对比较年轻,发生变异的年龄大约在5万年左右,M7与D单倍型类群发生变异的年龄大概在4万年左右,单倍型类群M7的溯祖年龄大概在四万年左右,单倍型类群F在海南黎族人群中的溯祖年龄为5.1964±1.9914万年,而在广西壮族人群中为5.3275±1.8913万年,这很可能说明黎族祖先的人群迁移历史是一个很长的过程。对M12单倍型类群年龄的估计可能作为海南黎族人群分化的时间。广西汉族、壮族及海南汉族、黎族线粒体单倍群年龄估算表。(2014.9.20) 中科院昆明动物所的研究人员搜集了来自缅甸的14个群体的845份样品,及已报道的缅甸及其周边115个群体的5,907个个体的线粒体DNA(mtDNA)数据,从遗传学角度证实了早期人类从非洲迁徙至东南亚后,除沿海岸线迁徙外,还从内陆路线从东南亚直接进入东亚内陆。缅甸人群中富集着大量的mtDNA基部世系,年龄在5~2万年前,缅甸人群和中国西南人群共享了一些单倍型类群。搜索了47,873个亚洲人mtDNA数据发现,一些单倍型类群仅存在于缅甸和中国西南。基于mtDNA全序列的系统地理分析表明,这些遗传联系一部分是近期基因交流的结果(如M24、M90、M91),而另一部分则很可能缘于从缅甸到中国西南的古老迁徙(如M54、M55、M84),时间可追溯到2.5~1万年前。>>>、Scientific Reports。(2015.4.1) 由张亚平院士指导的研究生彭旻晟等对海南黎族3个族群285份样本的mtDNA遗传多样性进行了分析,结合以前已发表的数据,经更为细致的研究发现,mtDNA单倍型类群M12、M7e和M7c1*是揭示人类早期移居海南岛的候选标记,而基于mtDNA全序列测定的系统地理学分析表明,早期移民发生在大约7千至2万7千年前。>>>(2011.2.19) 东亚特有mtDNA M9a'b是一个追溯盛冰期以来人类迁徙活动的有效候选遗传标记。博士研究生彭旻晟等在导师的指导下,对该单倍群进行了详尽分析,结果显示,中国南部及东南亚是盛冰期之后人类迁徙的一个策源地。在遗传学上揭示了一条源自中国南部向西延展到印度东北部和喜马拉雅以南地区的内陆史前迁徙路线,>>>。(2011.1.12) 孔庆鹏等:中国84个族群6093例mtDNA样本数据显示,华南为东亚人群最早分化之地;新鉴别的基部单倍群如M71、M74、M75、M76、N10、N11等很可能代表了最初进入华南的现代人祖先的母系遗存,而非来自邻区东南亚的近期基因流,考虑到这些单倍群主要集中于华南,提示华南很可能是现代人类祖先进入东亚后的主要聚居及分化地。(2010.8.20) 2025年02月14日 一条Y染色体 李於菟网络文章:“例如诞生于东南亚半岛的父系基因K2,他的后代占到了全球男性人口的大约50%。后代分为两个大支,分别是K2a和K2b。”“K2a的现存后代主要分布在东亚、东南亚地区,在大约一万年前分支N北支的部分族群,不远万里迁徙去了北欧芬兰。K2b的现存后代主要分布在欧洲、中西亚,大洋洲,伴随着西方殖民兴起,数百年内他的后代又大规模进入美洲和澳洲。”“根据对多个至少在4万年前,从东南亚地区扩散到东亚和欧洲古人DNA数据的研究,史前古人迁徙的复杂性,远超过我们的想象。”“不论是K2b往东亚迁徙,还是K2a和F—F27277往欧洲的迁徙,无一例外都以失败告终。(彻底团灭,没有后代)”“以前一直被忽略的真相,大约4万年前全球可能发生了某种剧烈灾难,不仅其他人类亚种都灭绝了,我们智人的祖先也险些走到灭绝的边缘。但东南亚那一块温暖隐蔽之地,终究还是为我们智人保留了火种,让我们得以繁衍至今,并且统治了地球。”(古DNA证实早在四万年前,现今东亚人的“祖先”就已经进入了欧洲) |


{kind=link}
{kind=link}