![]()
|
|
|
�� |
�л�Դ����Ҫ <15>
<14> <13> <12> <11> <10> <��> <��> <��>
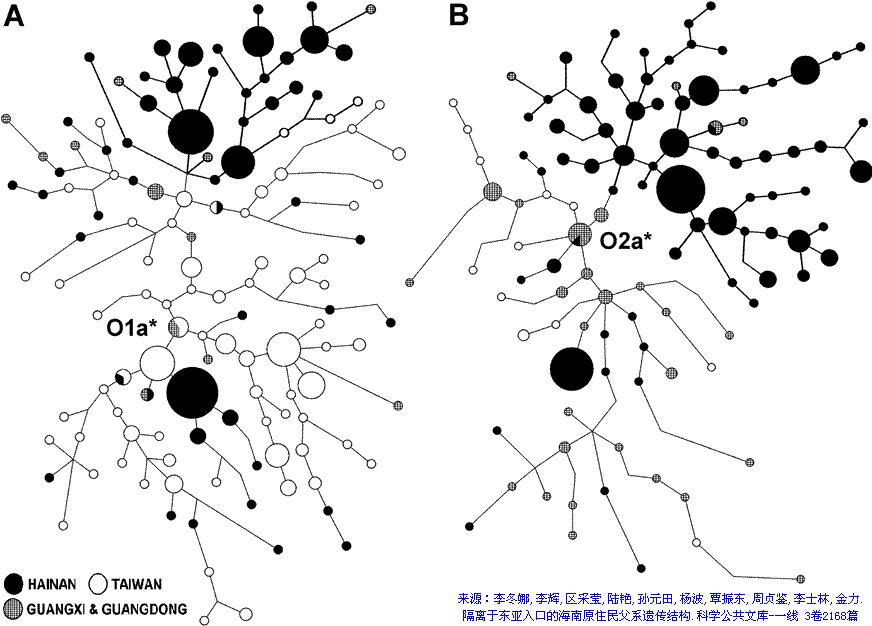
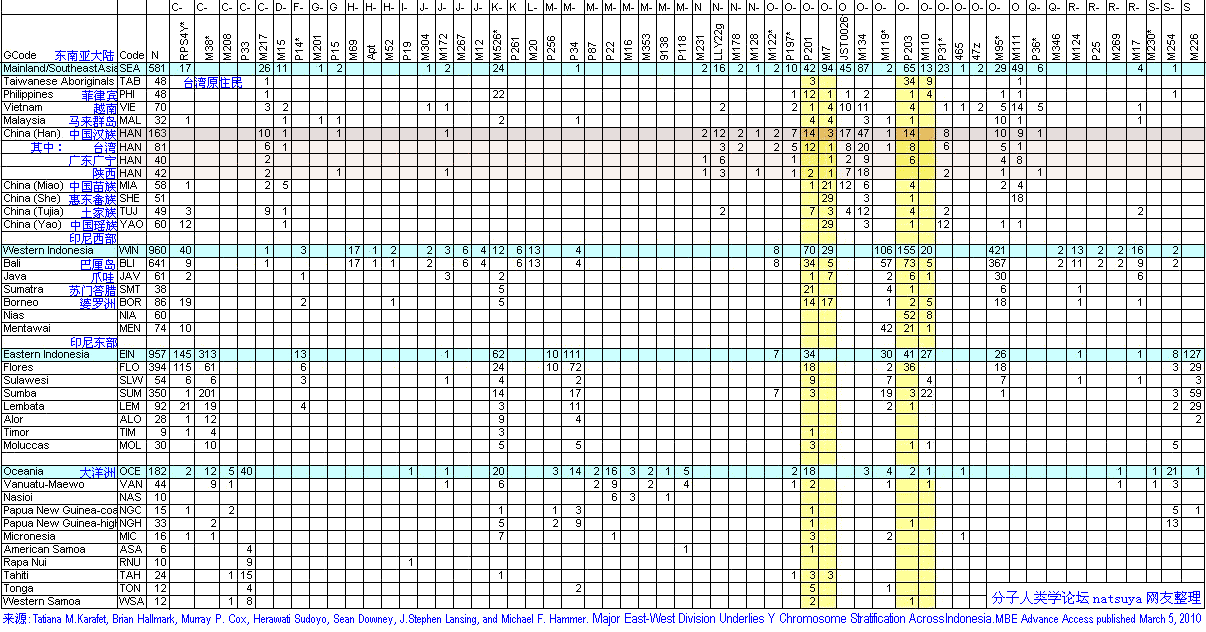
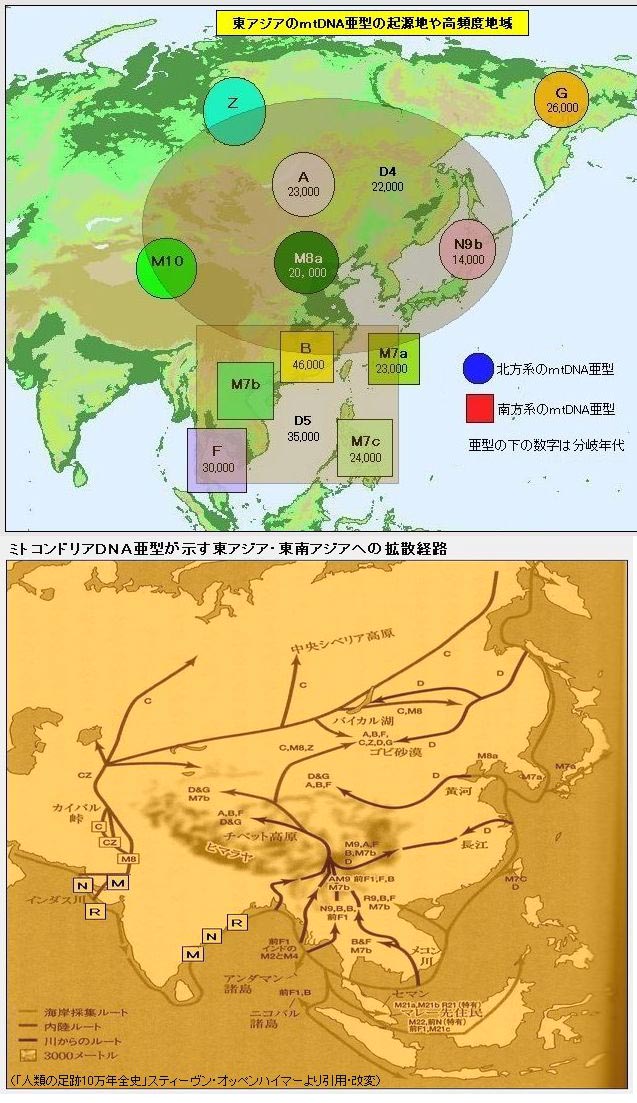
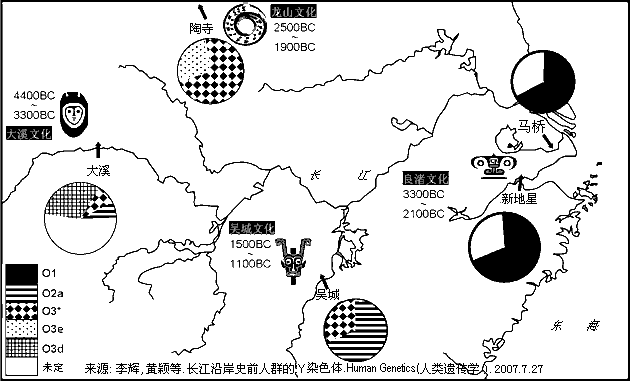
<��> <��> <��> <��> <��> <��> ��ѧ���� Ī�粨 2010 ��������.����.���� ���ϡ����ϰ뵺������֮��ĸ�ϵ��������ϵ����.D���붫������(5�����ǰ)�����ھ۾�������һ����N��Լ2����ǰ�����ϵ����������ϰ뵺���ϵ���D��N��ϡ�٣���˵����ʯ�����ڼ���ʯ��ʱ���������ڴ����������ϰ뵺�����Ǩ�㣬�ڼ��������塢����֮���ɽ�����������δ�������ϰ뵺�����벽����.������ʯ������ʯ�����ֵ�����������ϰ뵺�磬�Ļ���ͬ��Ҳ������֫�ᣬ����Ƿȱ��ʯ������ʯ����������(����ǧ��ǰ)����˵�����ϰ뵺��ʯ���Ļ���Դͷ���������ϣ�����������ʯ���Ļ������ϰ뵺�����������Ϸ���Ĵ����������ϰ뵺�����Ϸ����Ǩ��·��Ӧ�����ڡ���.������Ⱥ�ϸ�Ƶ��O*����O2*-P31��O2*�����ϰ뵺���٣����ϵ�������Ҳδ����ӡ�ȵ��ɴ���Ⱥ��O2��O2a�ĵ�����Ӧ�����ϣ�Ȼ�����������ϰ뵺����Ȼ����ϵ��ϸ���������ϵijɷ�O2a��Դ�����ϣ��������O1a(P203*)��O3(M117��M7)��ӦԴ�����ϡ���.�ݲ������������ݣ��ϸ���O1a������Ϊ5/689(0.7%)��̩��O1a����Ϊ17/365(4.65%)���й���̨O1a����Ϊ144/680(21.1%)���ϸ��������ϰ뵺�������Ⱥ��O1a�����ǣ���Ȼ����O1a��Դͷ���ڣ��ٽ����ȵȵ�O1a����ṹͼ��̨��ԭס����ӽ���������ģ���ΪO1a��ԴͷӦ��������������һ�������������㣬��26.6%���й���̨���Ļ���ͬ����73.4%���ϸ����������γ������ϰ뵺��̨-̩�����塣����Խ�����Ͻ���ѹ���ϵ�����ж϶�̨��Ⱥ��ǨӦ������ս�������ڣ�����������ѧ�Ƕȷ�����Ǩ������ڹ�Ԫǰ��������ǰ������(1989)�����ڵ�̩��������ܾ��ڰ뵺�����غ����غ�ʱ���˼�ѹ��Ǩ���ϲ�����½����.ͬ���������Ƶ����ݣ�M7���ϸ��ޡ�̩�м��������нϸ�Ƶ�ʣ����������ǵ���������Ⱥ��������������У����϶�̨987������������1��M7����˵�����϶�̨���е�O3ϵͳ������Դ�뵺�ϸ��ޣ�Ҳ������Դ������ľ�������������ԭס������Ա������壬ͬʱ�жϣ����������������Ǩ���뻪�϶�̨�Ӵ�Ӧ�������ڵ��¡���.̨��ԭס���м���O3a3-P201*Ҳ��O2a1-M88���ϵ���P201*Դ������ǧ��ǰ��̨��(��Karafet)������ʯ�����ڵ��ϵ�����P201*Ҳ��M88����˵��P201*��M88��ʢ���ڽ�������ƽ������ǰ���ü���̨��ȴ�����������ϰ뵺Ǩ���ϵ���P201*��M88����Դ�ز����������ϰ뵺��Ӧ�����ϡ���.ӡ���ɴ���ȺO2a��Ƶ���乲������Լ1.5����(��baiyueren;����2010������,ӡ����������O2a�Ĺ�������Ϊ17��28ka)������δ��M88��Ҳδ����O3(M117��M7)���ٽ��ʢ���ڽ�����������ж��ϸ�����Ⱥ���е�M88��M117��M7�������ϰ뵺�����Ӧ��������ǰ������ʯ��ʱ���൱�����ͼ����>>>��������ݣ������������,�IJ���natsuya��������չ��>>>��(70) ��������.����.�� ���ļ沢���ĵ�ĸϵ����Ⱥ����(�����Ƕ�56��57��62�����벹�䣬������ȺMϵ�Ժӱ�ĸϵ�ĸ�������أ����Ժ��ϡ�ɽ���̶Ƚ���)���˾�ǧ��ǰ���ڻƺ����ε�μˮ��������ԭʼ����O3����Ը�ϵN��Q1�������������ʹ����N��Q1���뵽������Ⱥ���У�ͬʱ��ԭ�ȸ�ϵN��Q1��Ů��Mϵ�������뵽������Ⱥ���У���������������������Ϊ������Ⱥĸϵ�����壬��������ȺO3��ԭ��Nϵ��ѹ���١� ��������.9.��7 ��̸������Ⱥ���嵥��ȺO3��Ǩ��·�ߣ�������Ⱥ���л�����������ҪԴͷ������̽Դ�¹��ش���Ū�������л�Դ���Ĺؼ���Ŀǰ�Ļ�����ʶ�У���.������Ⱥ��Ҫ����ũ��ҵ����ǰ�����Ļ��������Ļ�����ԭ��ɽ�Ļ������뱱��N2��N3��R1��Q1��C3��Ⱥͬʱ�ڵ����Բɼ��Ļ����Բ�ͬ����.������Ⱥ����(����ͷ��)�������������Ż������ͣ���Դͷ�ڻ��ϣ���.������Ⱥ��ϵ���������Ϸ���Դ��O3ϵ�������б�����Դ(����)��N*��N1��Q1��C3*����Ⱥ��ĸϵ��MϵΪ���塣��Ҫ����ǣ�������Ⱥ���嵥��ȺO3��Ǩ��·����������-�Ĵ���������-�����ߣ�˵������-�Ĵ������ߡ������������ǣ�ĸϵ��Mϵ��Ƶ����ϵ����һ��������N�������Խ��͵������ǣ�����ͬ·���ĸ�ϵN����Ϊʲô���ڱ��������γ�ũ��������ȴ��չΪ���������ߣ�O3Ϊʲô����N����һ��Ǩ�㵽���Dz��γ�O3ϵ�������壿���У����ϡ��Ĵ�һ�߲�δ�����κ���ʯ������ũ���ż���˵����-���ϡ����ߡ������������ǣ�������Ⱥ�����嵥��Ⱥ��O3ϵ����ʯ�����ھʹ���ũ��ҵ�����붫���Ķ��ļ����ϲ��ľ�����ͬ��ͬʱ���붫�ġ�����һ�������������������Ż������ͣ��Դ�˵���������ǣ�������Ⱥĸϵ��������Mϵ�������ġ�������ĸϵ���нϸ߱�����Nϵ���ɼ�����Ƶ��Mϵֻ�ܴӡ����ߡ������Ҵ𰸡� ��������.��.���� ̨��ԭס���O3*Դ�Ժη�������Եȡ������������У�O3*����������ռ27.3%����������ռ21.4%��������ռ14.3%����Ͷ���˼�̨�����ϸ�ռ9.1%��Karafet��̨��ԭס��48������3��O3��ΪO3a3-P201*��natsuya���Ѽ�һ��δ����������O3a3-P201*������ռ35.9%��������ռ17.4%�����ɱ��͵�����ռ12.5%�����ɱ���(���͵�������)ռ19.7%�����̨��ԭס���O3a3-P201*��Դ��ʢ���ڵĻ����غ�����ΪO3�����ڹ㶫�����ṩ����֤����Karafetͨ���Ա�O3a3-P201*��STRֵ���֣�̨��-���ɱ�-ӡ��-�����ޱ˴�����������½���൱Զ������Ϊ̨������ϵ���������(��ʯ��ʱ��)ͬԴ��������Ӧ��������½�ֿ��ˣ�����̨����ϵ�Ӧ��ʢ���ڶ�������ʯ��ʱ���������ϵ���O3���½�ķ�����磬2008����������Ҳ�����ͬ���Ŀ�������>>>(26)�����ǣ�����̨��ԭס��ĸϵ��18%�ϵ���Դ��Eϵ�������ų�O3a3-P201*��ΪĸϵE����Ӧ�ĸ�ϵ���ϵ����ɱ��������̨�塣��ϵ��O3�ȵ����ϵ��ģ���һЩ�����Ǵ�½û�л��еĵ���Ⱥ����C2-M38*��K-M526*��������C-RPS4Y*����½�У�����Ƶ�öࣻ����ϵ���O3Դ�����ϰ뵺����������е�Ӧ��O2a-M95*��ӡ��������ȺO2a-M95*������43.8%(421/960)����O3a3-P201*��Ƶ�öࡣ����̨��ԭס���O3a3-P201*�����ϵ�����������ӡ�ᡢ���ɱ���·;ңԶ���������ӣ���;�ض������C2-M38*��K-M526*��C-RPS4Y*��,��һ����Ⱦ�����������ܣ�����O2a-M95*����ҲӦ��O3a3-P201*�ߵöࡣ��̨��ԭס��ȴû����C2-M38*��K-M526*��C-RPS4Y*����O2a-M95*Ƶ�ʱ�O3a3-P201*���͡���Щ˵����O3a3-P201*Դ���ϵ������ǵò���֧�ֵģ�̨��ĸϵ�е�EӦ��̨��ԭס���������̴ӷ��ɱ��ȵش������ģ�Ư���������������Ϊ���壬����ȡ�óɹ�������ϵ�Ů��Ҳ�ǺϺ������ġ����ϣ�̨��ԭס���O3a3-P201*��O1a1-P203*Ӧ��Դ��ʢ���ڵĻ��ϣ���O1a2-M110��Լ��2����ǰ������̨�壬ͬʱ��O3�ĵ�����Ӧ�ڹ㶫����һ�����������ϡ�(������������̨Ͻ��������2��8.2��7.5kaB.P�Ĺ�mtDNAΪE��R9���������ڶɺ��������ӦΪ�غ�С��ԭס��E��ԴͷӦ��̨��������ϵ���E��M9������M9a���еķ�֧��������2.7����ǰ���������ϵ�������������ͣ���½��������������E��Դͷ���ڶ����غ���E�ܿ����븸ϵM110�������Ƶ�Ǩ��·���������˿�������̨�庣Ͽ��ƽ���������ڷ�յ�С�������´�������ľ�ʯ������������֫������˵������������Խ������ԨԴ��(67)
��������.��.���� �ϵ�Դ����Լ6����ǰ������ڣ���ƽ��ϵͣ�C-M130*��C2-M38*��K-M526*�������ϰ뵺������Ǩ���ϵ����ܵ���ĵ������Ƕ�ռ�ˣ�����ĩ��ʢ���ڣ�Oϵ��O2a-M95*��O1a-M119*��O1a1-P203*��O3-M122*�����ϰ뵺������Ǩ��������ӡ������ϵ����ھ��������������ӡ��Ⱥ���������ޣ�Oϵ������ӡ���ʯ�����ھ�������ӡ��960��������Oϵ�����615��ռ64.0%(����Ϊ������Ǩ��)��ʢ���ڽ�������ƽ�������������ϵ��������½����������һֱ��Ϯ�ž�ʯ���Ļ�����½���ڵ����ĵ�������O2a1-M111��O3a4-002611��O3a3c-M134��O3a3c1-M117��Ҳ���ܽ����ϵ��������ϰ뵺��������Ⱥ���У�O2a-M95*��O2a1-M111(M88)���У���ӡ��447��O2a��ȫ��M95*����Ҳǡ��˵����O2a-M95*�ھ�ʯ��������������������6ǧ����ǰ�������غ��İ�Խ�������������¼�Ư����̨�壬���ǰ���ʯ���Ļ�������̨�壬ʹ�����ˡ������˺���һ���ӴӾ�ʯ��ʱ�����ɵ���ʯ��ʱ���������д�����Ļ�����Լ��ǧ��ǰ����½�ĵ���ũҵ����鮼���Ҳ����̨�壬�������¼�����ԭʼ̨���˿�ʼ����ɱ��ϵ��������Ǩ�㣬��Լֻ����500��ʱ��ʹӱ����ϡ����������ϵ��Ӿ�ʯ���Ļ�����Ϊ��ʯ���Ļ�������������ͬ��Ϊ�ϵ���ϵ���������γɡ���̨�����ϵ�������ʯ���Ļ�������ϵ����ȺΪO1a1-P203*��O1a2-M110��O3a3-P201*���伸�������˵����ϵ�O1a2-M110��O3a3-P201*��ȫ��������O1a1-P203*������ӡ��960�������У�Դ��̨���������������165��ռ17.2%����С������̨�������ܹ��ڼ�����������ͬ���˴�������ϵ���������ȫ��������ʯ���Ļ��ľ������ƣ�����ռ��ӡ�������43.8%(421/960)��O2a-M95*���Ǿ�ʯ�����ڵ��ϵ�������������ʯ��ʱ��Դ�����ϰ뵺���������壬�ϵ������ͬ������������ô��Χ��óɹ���(��زο���ͼ��>>>������>>>���������������������ǿ��ż�����ģʽ��������ۣ��ϵ���M119*��M95*��M122*Ӧ��ĩ��ʢ��������) ��������.��.���� Y-SNP����ȺC3*Ƶ�ʱ�����������Ϸ������ԭ��������������ȺCϵƵ�ʷֲ������������罭�պ��塢�Ϻ����塢���պ��塢���塢ˮ�塢�����塢����������Ȳ�����Ⱥ�⣬�й�CϵƵ�ʷֲ�����ǣ��Ϸ��������壼�Ϸ����壼�������壼�����������壬��С�Ӽ����ٷֵ㵽ʮ�����ٷֵ��ٵ���ʮ���ٷֵ㣬�����1:2:4:8������������������C3*����������Ϸ���������������弰�Ϸ�����������Ҫ��C3*���ֲ���C*������û��C3c����������������ΪC3*��C3c������Ⱥ�л��нϴ������N2��N3��ͬ�Ǻ��壬Ϊʲô������C3*Ƶ�ʸ����Ϸ����ѵ���ʷ�ڱ��������������롰�������Ľ������ʷ�����������غ�ʱ��ū�ĺ���������ʱ�ʱ���ĺ��������������塢Ů���塢�ɹ��塢������ռ���������������ԭ�������弸��������C3c��N2��N3�����ԡ����������Ʋ��Ƿģ�Ҳ����˵�������ϸ�Ƶ��C3*������Դ�Ժ��˵����룬ֻ�ܽ���Ϊ�����ڱ����γ�ʱ��ԭ���ͻ���нϸ߱�����C3*���Ϸ�����C3*Ƶ�ʽϵ�����Ϊ��������ʱ�����C3*Ƶ�ʸ��͵��Ϸ�ԭס����ϡ�͵Ľ�����ۺ���ǰ�ķ��������ڻ�����Ⱥ�Ĺ��ɴ���ǣ�60%O3��15%C3*��10%N*��5%O2*��5%Q1��5%����(��D)���ڶԶ��ļ�������ͬ���ںϹ��̣�O3�������ߣ�N*��Q1�������ͣ���C3*�仯����������������Ϸ�ԭס��Ⱥ��ϳ��Ϸ�������̣�N*��Q1��C3*����������O2*������ ��������.��.���� O1a2-M110��Ⱥ�ĵ�-½Ǩ�㣺M110�ǰ�ԽM119*�����η�֧����̨��ԭס���г��ֽϸ�Ƶ����������Ϻ����š����ԭס���Լ��ϵ�ӡ�����Ⱥ���������������������������M110Ӧ�ǽ����Խ�����������š����ԭס��ͬԴ��̨�塢�Ϻ���ӡ��ȵص�M110��Ⱥ���ңԶ�����д����������֮�便������ʲô����ͬԴ��ϵ��˭��Դͷ�����Ǩ�㣿�Ƿ���ھ�ʯ�����ڻ�����ʯ��ʱ����
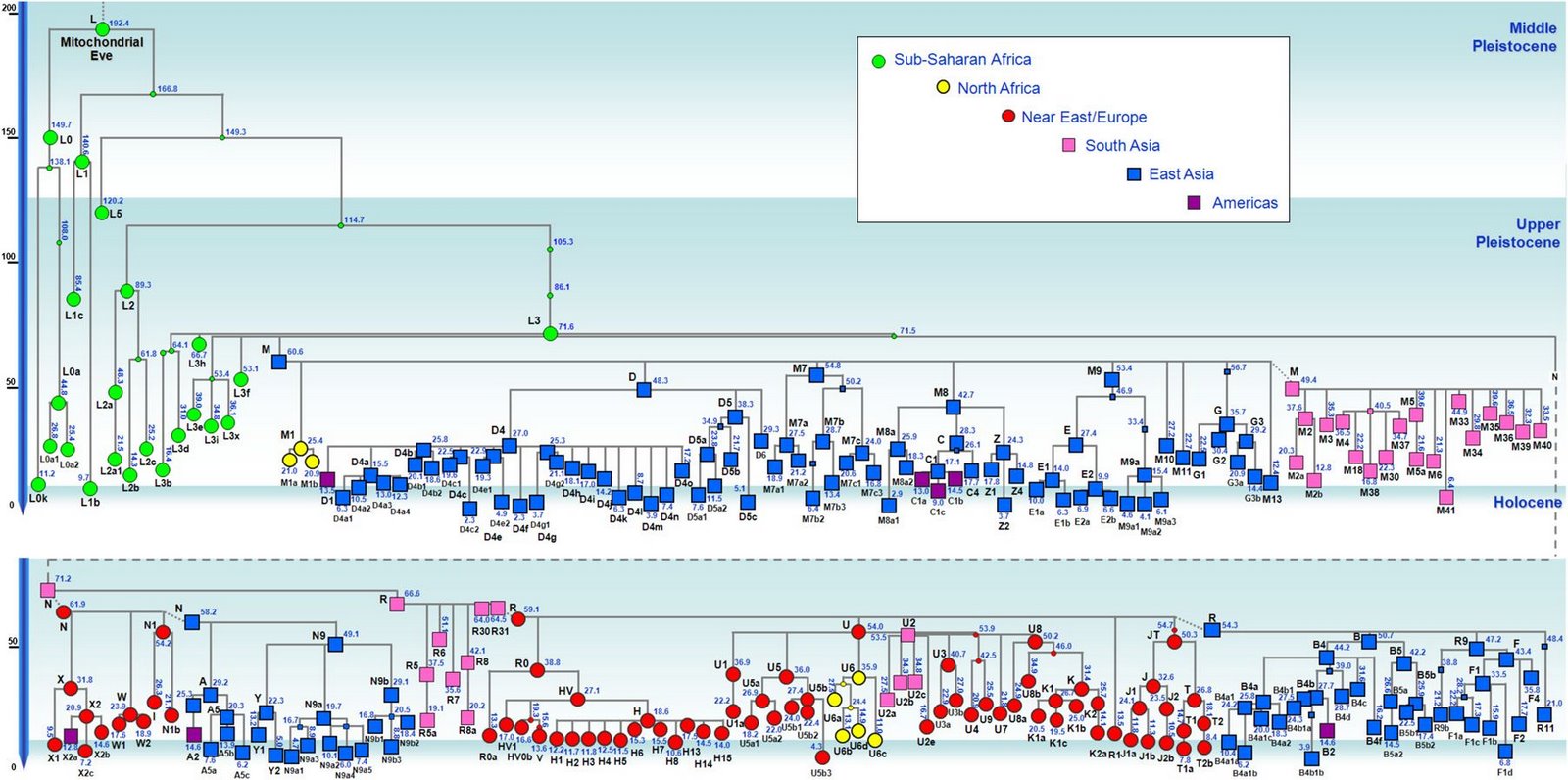
��������.��.�� ������Ⱥ�еĸ�ϵQ1-M120��Q1�ڶ�����Ⱥ��Ȼ��Ƶ�����ֲ���㣬��ΪΨһԴ�����ǵĸ�ϵ�����ͣ���ʹ���������棬�ѵ���ˮ����ʯҶ�Ļ��ĺ��ˣ���Ϊ����ϸʯ���Ļ��Ĵ����ߣ�����Դ����ū���ʱ��ĺ�����Q1��ʲô�����ʲô·������ǵģ����Ǹ����������������������? ��������.��.���� ������Ⱥ��Դͷ������ģʽ����������ĸϵ��MϵΪ�����������ž��������������ѧѧ����28����4��(2009.11)�������������˹ǵ�mtDNA�����������������̫��ɽ������������4ǧ��ǰĸϵ������MϵΪ����ͬʱ�����������ɽǰս��Ĺ��ɽ���ȵص�Nϵ���ݣ���Ϊ������Ĺ��Nϵ�Ǻ�ӦΪ��������ս�ܹ��ķ�²��������Ⱥ�Զ��ĵ���Ⱥ�ļ沢���ںϴ����൱��ĸϵMϵ���������������븲�ǹ�����������֪�����ǣ�������Ⱥ��Դͷ�����ĸϵMϵ������ʲô��ʽ��Ϊ������Ⱥĸϵ������ģ���Ҳ������̳�����۵Ľ��㣬�����۵���Ϊ�����ĵ�Դͷ����Ǽ��˵2�����ǰO3��ϵ�е�O3e-M134(��M117)Я����ĸϵMϵ��������̣�Ǩ��������������Ȼ��ֳ�������Ⱥ������֧ϵ������O3*(M122��M324)֧ϵ�������س������£�Ȼ������·�ߣ��ں��ϡ�ɽ���ȵ��γɶ��ļ��ŵȣ����������Ļ��������ţ��붫�ĵ���Ⱥ�����沢�ںϣ��γɺ��壻�����Ҽ����Ϊ��ֻ������-����-��ԭ·����������O3e-M134�������ߣ�Ҳ������һ��������M134ԭʼ����Ⱥ�䣬μˮ����Ļ��IJ���ֻ�����ں���O3ϵ��һ����֧��M134��������O3ϵͳ��Ⱥ�е�һԱ���ڻ�����Ⱥ�����Ź��̣�M134(��M117)��M122(��M324)������ͬ�������������ŵġ� ��������.��.�����ǹ���O3a3c1*-M117(O3e1-M117)�����⣺��ʯ��ȶ�λ���ߵ���������ͼ���У�M117�����Ǻ���������Ա(M117��M134��M324��M122��M9)֮һ��Ҳ�㷺�طֲ�����������֮�У�����Ƶ�ʸ��ߣ���M324��M122������ͬ��M117��M134����������姡���Խ���Ϸ�ԭס��Ⱥ���зֲ�����δ��������̨�弰���ϵȺ���ԭס��Ⱥ֮����M117��M134�ܿ�������ʢ���ڽ�������ƽ�������Ժ�����ŵġ���M117��M134��STR�ṹͼ�ϣ��Ϸ�ԭס��Ⱥ(��������姡���Խ)������ϵ�ĸ���(ͼ�к�ɫ�ϴ�Բ��)��˵�������������Ϸ�����ʾ��ͼ>>>��M117��ԭʼ�ͼ���������������Ⱥ(������)���������ʼ��ؿ����������������ʼ��ؿ����ڹ���(O*�ĸ�Ƶ���ڹ���)������M117�ĵ�������M134��Զ�����������������ϡ���һ�����ڴ�������У�δϸ�ֵ�M134�����M117��M117������Ҳ�ܴ�����M134(xM117)��ƽ���²�M117�ķֻ����Ӧ��M134�ӽ�����M134�����ñ㻯�ֳ�M117��M134��M117������O3ϵǨ�㱱��ǰ�Ѿ�����������ǰ����ָʢ���ں�ƽ����������Լ��������������ʯ�������(1.2����ǰ)���ж�M134��M117Ӧ������1.5��1.3����ǰ�������ĵ���Ӧ����M7��1.2����ǰ��M134��M117�л���M7ϵͳ�ľ�����Ⱥ����M7ȴδ�ܻ���M134��M117ϵͳ�ı�����Ⱥ֮�С�֮ǰ�����ڶ�M117��������������ص����У���Դ�������������Ǻ���Դ���ķ������������ص�ƫ����٣��Ϸ�����������ߺ����ٸ��˵ȵ�M134��M117��һ��Դ�ڱ������ˣ�Ҳ��һ��Դ�ڶ��ģ������������Ϸ�ԭס�����O3ϵͳ�����е�Ƶ��Q1�����Ϸ�ԭס��Ⱥ��O3ϵͳ����û��Q1���֡�(61) |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}