![]()
|
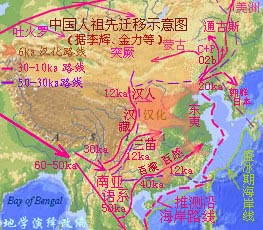
|
|
�� |
�л�Դ����Ҫ <15>
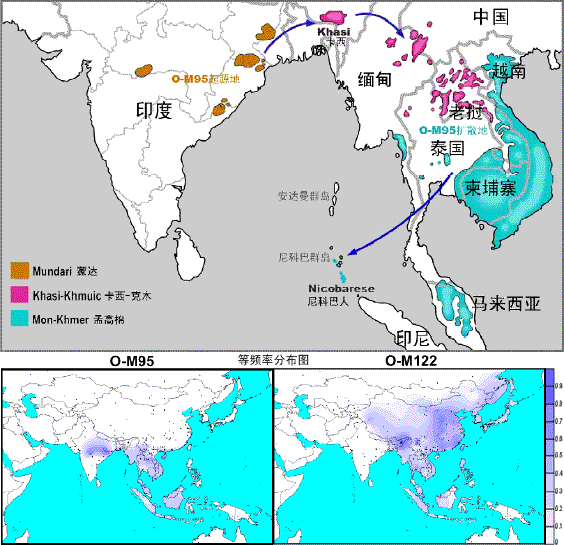
<14> <13> <12> <11> <10> <��> <��> <��> <��> <��> <��> <��> <��> <��> ��ѧ���� Ī�粨 2005��2008 ��������.��.���� O-M95���п�����Դ��ӡ���ɴ���Ⱥ�����ȣ�������ڡ�YȾɫ��֤����ʾ������ϵ��Ⱥ�й�ͬ�ĸ�ϵ��Դ��(��ͼ)��ָ����.ӡ�ȴδ�½���ھ�ס���ĸ���Ҫ����Ⱥ��������ϵ������ά����ϵ��ӡŷ��ϵ�Ͳ������壬����������ϵ����Ϊ��ӡ��������ϵ������ϵģ�ͨ���Ƚ�������ϵ������������(�ɴ����-��ľ���ϸ���)��Ⱥ��YȾɫ������ԣ�����O-M95������������ϵ��Ⱥ������Ҫ�ĵ���Ⱥ������ӡ���ɴ�����Ⱥ��STR��������ߣ��Ӷ��ó�����ȺO-M95Լ��65,000��ǰ(95%CI:25,442��132,230)��Դ���ɴ�����Ⱥ�����Ľ��ۡ������ǵ�������ϵ��Ⱥ�Ǵ������ɴ�������Ⱥ��������ӡ�ȶ�������Ǩ��������������������ѧ֤��һ�£�������Ϊ�ɴ�������������ϵ�������������ӵ�����(6����ǰ�������п���Ӱ�쵽������?)��������ָ��һ������Ⱥ���п�����Դ�ĵط����������������ƶϣ��ߵ�Ƶ�ʺߵĶ����ԡ��ڶ����ǵ�������ϵ��Ⱥ�У�O-M95�����Ƶ��Ϊ35%�����ɴ�������Ⱥ�����������ܴ�(35��109),O-M95��Ƶ��ƽ��Ϊ63%�������Աȶ����ǵ�������ϵ��Ⱥ��Ҫ�ߡ������о��ó���Ǩ��·��Ϊ���ɴ������(65,000YBP)�Dӡ�ȶ���(57,000YBP)�D������(8,000YBP)�D��ư�Ⱥ��(17,000YBP)(����ܳ�ǰ���ǵ���ȥ?)���½���һ���о��ᵽ��O-M119�����ԼΪ20,000YBP�������˵���ڶ�����O-M95��Ҫ�ֲ���½��O-M119��Ҫ�غ����߷ֲ���O-M119����Ǩ�㷢���ý�Ϊ��������ˣ���ȷ��O-M95���Ҫ��O-M119���ϵöࡣ��.���ɴﲿ���붫���ǵ���Ⱥ��û�����Ƶ�ĸϵ������ϵ�����Ƿ�����1147�������������˴��ɴﲿ���Լ�����Ⱥ�壬��ȫû�з��ֶ��������mtDNA����Ⱥ����ʾӡ�Ⱦ���������ϵ�����������в�ͬ��ĸϵ�Ŵ���ʷ����һ�������ڹ�ͬ�ĸ�ϵ�Ŵ��Dz�ͬ�ġ���Ȼ������ϵ�������й�ͬ�ĵ���ȺO-M95��mtDNAû�з��������������ô��һ������ռ������ǰ�������ǵ�Ǩ����Ǽ��п��ܵ�(Ϊʲô��������������Ϊ���Ĺ�����Խ��Ǩ��ӡ��?)��������Ҫ���ǣ�O-M122����Ϊ�Ƕ�������Ⱥ����������Ⱥ�����ɴﲿ��������ȫ�����ڡ����κη�������ʼ�Զ����ǵ�Ǩ�㶼��Ҫ��O-M122Ϊ�����ġ�����Ⱥ����õ�O-M122�������15,000��60,000��ǰ֮�䣬�������ǵ�O-M95������Ϊ8,000��ǰ�����ӡ�ȵ�������ϵ��Ⱥȷʵ���Զ����ǣ���ô����Ӧ������O-M122��������Ϊ���ֽ��ɴ��˵������Ǵ�ӡ��Ǩ�㵽�˶����ǣ��������෴(ʲô?�ⲻ����˵�����Ƕ�����������û��M122��,������ԽǨ��ӡ��֮��,M122�Ŵ��������µ���?)��O-M95���п�����Դ���ֽ��ɴ���Ⱥ�����ȣ�����Я������Ǩ�㶫���ǡ�(ժ��2007�꡶�ִ�����ѧͨѶ����һ��.��̬����.����.��\��) ��������.����.�� ��¡����ַ���������ַͬ�о�������˵������Դ��ͬ���������ɺ��������¡����ַ(8.2��7.4kaB.P)��30�������صľ���Ĺ�ᣬ�����ľ���������ӳ����һ��Ũ�ص��ڽ̼�����ʽ�����߱��������ڣ���Ϊ���߳�ݡ�����Ķ�����������Ϫ�ص��������ַ(7.0��3.5kaB.P),�����˰˾߱�������������˹ǣ���Ϊ��֫�ᣬ�������ھ����ڡ�����ַ�����ϵر���ȴ����ͬ�����ף��ܿ�������Ϊ���Ǿ�����ͬ����Դ����ʯ���硢���ڵ���֫����Ҫ�ֲ��ڹ���-Խ��һ����Ӧ����������ԽO2a�����ף������������Ӧ����ԽO2a�����ɴ��жϣ�������¡����ӦΪ��������ԽO2a������O2b(������O2*)����¡���˵����ȹ�����ʢ���ڽ�����(Լ1.5����ǰ)�뿪�������عź���ֱ���ϵ������ɺ�������ַ�����������������������ǡ��������������Լ���ũ������������������¡����Ӧ��Դ���Ϸ�����Զ�ŵķ��g���估ʼ���gΤ����ԨԴ������¡��-��ɽ�Ļ����ڿ������Ϸ���OϵΪ���������Ƶ��˺�ɽ�ڣ�����Ը�ϵN���˷���������ںϣ�ţ����ĸϵ��NϵΪ������ϵ��N*ռ���ƣ�������O������C*����(89)��(16) ��������.��.�� �����Ƿ���O2bӦԴ�ڱ�����-����һ�������ȣ�O2b��O2a�����ף���������Դ��D��C��P��O1��O3��Ҳ������ֱ�Ӵ�O2a�ֻ���ֻ����Դ������O2a��ԴͷO2-P31����2�����ǰ���й��Ϸ������ϰ뵺�����غ�����Ǩ�㼰��ɢ���̣���ʼ����������O2*�����ﶫ����ֻ���O2b���ڳ��ʰ뵺���ձ����ٷֻ���O2b1����Σ����ϵ������塢Խ��(Լ4.7%����)��ӡ��-�����ˡ���Ⱥ���ܿ�����������Ҳ��O2b����ΪO2b���Ϸ���Դ˵�ṩ�����ݡ���������ʼO2b(��O2*)���Ϸ�����������غ��ı�Ե��Ⱥ������ʢ���ڵĽ�������ƽ����������ռ���С��O2b��Ⱥ�����Ŷ�����Ǩ�㶫�������ʰ뵺��̨��ԭס����ǷȱO2b�ģ�˵��O2b����ʢ���ڽ�����ƽ����������غ������ϣ��ձ��˵�O2b��ҪΪO2b1����Ĭ���ձ���˫��Դ��һ����Ϊ��O2b1�������ġ���ǧ��ǰ�ij��ʰ뵺�ϲ����ձ���O2b����O2b1�����Ժ�Ž����ձ��ģ�������߱����۳��������������ڵ��¡�(15)(��Ͽ���֤��,��O2b1�������˽����ձ���ʱ��Ϊ���2300��ǰ) ��������.��.���� ��������������Ӧ���綫����D��5��2.5����ǰ�ļ���ڣ��綫���˱鼰���Ǵ�½�����������ڵ���ʱ�õͺ�ƽ�浽���ձ��������ڣ���ƽ���������ձ��е����½����������������ʱ����½���綫���˱���ɱ�����𣬶����д�������ձ�ȴ������ر�����������ʹ�����ܿ��ӵ�ԭ��½�綫���˵���Ӱ����ʢ���ں��ƾ��ձ��Ļơ�������Ѫͳ�����Կ������ձ�ԭס��YȾɫ����ҪΪ�綫����D��Ϻ���˾���100%��DѪͳ���ɴ��ƶϣ����Ǵ�½����������ҲӦ���綫����D�������綫����C������ĩ��ʢ���ڼ����������˵���ɱ���綫����DѪͳ�ڶ��Ǵ����Ⱥ�еı����൱ϡ�١��������綫����CѪͳ���Ǽ̳�ԭ�ػ��ԭ�ص��綫���˶������ܿ����Ǻ����ٴ��Ϸ�����(14) ��������.��.���� ����������ʯ������Լ��ʼ��5����ǰ���綫���˴��϶����鼰���ǣ��������������Լ��4��3����ǰ�����Ϸ������о�����Ϊ���綫����ʹ�õ������ھ�ʯ�����������˽���ʹ����ʯ������ʯ����������������Ϊ��Ѱ���ǵ��ټ����б�ҪŪ����ʯ������ʯ�����ص㡣��ϴ�ª�ľ�ʯ������ͬ����ʯ����һ��������ľ�������ʹ�õĽ�Ϊ�Ƚ���ʯ�����ߡ��Ϸ����ڷ绯����ǿ�ң����ʶ���Ӳ����ʯ������£����˶���úӴ��Ĺ���ʯӢɰ������ʯ��������ʯʯ�������Գ��ֿ������ӹ�ľ���õ�����ʯ�����и���Ƥ����õ�ĥ��ʯ�����Լ���������ľ������ʹ�õ�����ʯ��Ϊ��������ʯ��ʱ�����Գ���������ũҵ������ҵΪ��������ȣ���ʢ���ڣ��������ٷ���������ĥ����������ʯ�������Ǽ̳������ھ�ʯ����չΪϸʯ��������ɽ����¶���ڲɾ����Ķ�Ϊ��ʯ����ʯ��������ľ����ϵ�ϸʯ����������ʯ���ߡ�ʯéͷ֮��Ķ���������ȱʧ��ʯ�����Ʋ���5��1.2����ǰ��û�л����˵�������Ϊ�綫���˵ĵ��̡�������Ҳ������Ϊ����ʢ���ڵ�ϸʯ������ʯ�����������������Ŵ滹���綫�����Ŵ�����ȷ�����йػ����о���Ϊ�����˴�Լ��3��2.5����ǰ���뱱���� ��������.��.���� �й�ѧ�߶Ա����綫����ѪͳC��Դ�Ŀ���������߱������綫����ѪͳC��Ҫ�ֲ��ڽϸ�γ�ȵ��ɹŸ�ԭ���������ǡ����������ʰ뵺���ձ��ȱ��ǵ����������ɹ��弰����ͨ��˹��Ⱥ��ҪΪC3c+C3*���������弰��������ҪΪC3*���ձ���������C1�������Ϊ��4��2����ǰ���綫���������������鼰�������ǣ���Լ��ʼ������������ǰ�������˽����Ϸ��������綫���˽����˲�����Ů�����𣬴��Ϸ�Խ����Ů�����綫����Ѫͳ�����൱ϡ�����������������綫�����Ӵ��ڼ䲢û�з���ͬ���ںϣ����ڻ����������ţ�ͬ������������ڳ����Ա�������ֻ������ƫ�������н϶��ĸϵ�綫����Ѫͳ�������ں���֮�С���������֮�⣬�綫���˱����ϵ������˲�Ըȥ�ı���-��ʱ�ĺ�������������������ǰ���P*Ѫͳ�»����˵Ķ��㣬���綫���˷�����ͬ���ںϣ������������ֽ�һ�������ţ���ԭ�ذ�Ѫ�Ļ������ٴ��ں�Ϊ���ذ��ֵĻ����˱������֡��й�ѧ��ǿ�����ǣ����������綫����Ѫͳ��ϡ���DZ�����������Ľ�������ǵ��綫����Ѫͳ����ԭ�ػ��ԭ�صļ̳��ԣ����������ɵ��DZ����綫������Ҫ�������ڱ����ĺ������������������������ڵı���CѪͳ�Ǻ����ٴ����Ϸ�Ǩ���ȥ�ģ��便����Σ�������ȷ����(12)
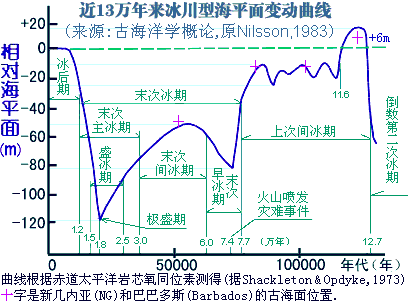
��������.��.�� ���ϵ������й������˶��綫���˵ĸ����������ϣ�������5��2.5����ǰ�ļ���ڡ�10��5����ǰ������ڣ�����2.5��1.5����ǰ��ʢ���ڣ��������ż����Ǵ��ڵģ���5����ǰ�������ˣ����ɳ����ַ7.9���ꡢ4.8���ꡢ2.2������������ֵ��3.9��3.5����ǰ����������Ƶ����ַ��4��1����ǰ����ε����ݰ�������ַ��1.6����ǰ��������ʯ����ַ�ȡ� ��������.��.�� ��������ѧ���о���Ϊ���й��˵�ֱϵ���ȴ�Լ��3��2.5����ǰ��Խ���������뱱���������ŷ����������5��2.5����ǰ�����������ڱ������ձ���ڵģ���ɽ��˷�������˼���ˮ�´��Ļ���ַ������ɽ�����ˡ�������������������ַ�����������ˡ�����ˮ�����Ļ��Ŵ桢���ɺ����ˡ�������ƽ�˼����Ӷ��Ļ��Ŵ桢���ְ�ͼ�˼��������˵ȡ�����Ŀ��������ܲ��꼼�����������ƣ�ȱ����ȷ��������ֵ���й���ѧԺ�ż�������������о������к硢ͬ���ĵ��˼�����ʥ-·��˹��ʢ�ٴ�ѧ��Trinkaus�����������˶��������ʯ���о����ó��������ʯ�ĵ������Ϊ4.2��3.85����ǰ�������о��ɹ�������4��3�ճ���ġ��������ҿ�ѧԺԺ�����ϡ�����Ϊ��������ĩ�μ���ڣ����������������˻�ģ���Է�������ѧ�Ǹ���ս���ѵ������������˺�����ʢ����ȫ���������˻����������Ϸ�����û����ԭ��ֱ�����ִ����ݻ���������ݼ�>>>��ѧ��(9) ��������.����.�� �������������ж�Ⱥ������ԴӦ����������������ɰ����������ڹź���ֱ���ϣ������Ͽ��Լ��1.4����ǰ���뱱�������Լ��1.2����ǰ���������ǡ���Զ����������1�����ǰ���������ɺ�����İ�̩��ϵ����������̩��ϵ��ź���֧������ͬһ֧ϵ��(·����ͼ��������ż�������������������)(8) ��������.��.���� YȾɫ��ͻ��ǰ����ɽ�˺�-������ͻ�������������M134ͻ���M117������5��6ka.BP��Ҳ����˵����M117�ĺ��˹�ͬ������ʱ�մ���̥����������ݿ��ţ�8ka.BP�ĺӱ���ɽ��ַ�ڴ��ؾ���10���ɼ��˿�֮�ڲ��ǻ����������6ka.BP�����������֣��Ƶۼ��������֮���ս������ɽ�˺�������������12ka.BP���������ൾ��9��8ka.BP���γɵ���ũҵ���������M119��Խ������8ka.BP���ڽ��������ͼ>>>(7) ��������.��.���� ʮ�־��ȣ�������Ⱥ������Ҳ���������������ǵ�����������Լ7.5����ǰ����ˣ����´ӷ����������ִ�������Լ��6����ǰ�������ǡ������ǣ����ص��������������������������������������̨����·Ǩ���й�(ͼ��������)��������2.5��1.5����ǰ��ʢ�����������ǽ��뱱���������������˼���ȫ����û��3��1����ǰ�������ڶ����ǵ��������Ȳŷ���·�������ǽ���(�Ϻ���)�������������ⳡ��������ɱ�Ļ���3��2����ǰ�Ĵų����Ե�ת�¼���WT��.�������ţ�WT��.�е㻬��-����ǵ��ϰ������������˶ѡ���ơ�!(6) ��������.��.���� ĩ�α���֮����������������죬����������������������˸�Ȧ��ʢ�����ֻص����Ϸ�(���ų�����ඳ���ڱ���)��ʢ���ڽ�����ȫ�������̣��ͷ��绢����ƽ���¡��̲����ޡ�>>>(5) ��������.��.���� ���˲�ͬ��ֲ��Ҫԭ�����������ǹ������Բɼ�����ʽ�������ת���ˣ��캮�ض������Dz����ܻ����ڸ�γ���������������������ů���棬����������Ǩ��Ӧ�൱Ƶ����Ѹ�٣��������ܺã�˵����������Ҳ�����ϱ������������ء����ڸ�γ�ȹ��������ʧ������������Ľ����������Ϊ��������(4) ��������.��.���� ��������ѧ��Ϊ���й�10��5����ǰ�Ļ�ʯ��ȱʧ�ģ��������˲�û���ִ����ݻ��������ڷ����ִ�������Լ��6����ǰ�����Ϸ�����3����ǰ�Ž��뱱����Ȼ���������Ա����5��3����ǰ���������˻�ʯ�����Ժδ��أ����������YȾɫ��Ǩ�������������������Ŵ�ķֲ�Ҳ���ϲ�������������Ϊʲô�����Ų���������еģ�����������˵����ȥ������˵��Щ���ϼһ��û���к����δ��Ҳ̫����ˣ�(3) ��������.��.���� ������Լ��6����ǰ�Զ����ǽ��뵽���ϵ�����3.0��2.5����ǰ���Ա��ı�����ɢ��������ɢ���ڸ��صIJ���Ӧ��չ���⣬��Ⱥ�ǻἯ��ϳ�һ���л�Ⱥ�塣Ȼ��������׳��������ֻ��Ǩ�㵽��������һС���ˣ�Լ��ʼ����ǧ��ǰ���������ǵIJ���׳�����������������ϣ�ͬ�����̲����������������䣬Ȼ���γ���һ�������ǡ���һͳ�����л�����(�����˿�Լռ92%)������ͼ��ʾͼ���ѵ�������ǧ��ǰ�ij������κ�ķ���Ļ����ƺ�����������-�����Ļ��ȶ��������ͬ���������ƺ��Ǹ�����ֵ����⣿(2) ��������.��.���� ����仯һֱ�ڡ���д�����й��Ŵ�ʷ����Ԫ1230��ǰ����γ����ҹ�����ת�䣬���볤�������߰���ĺ����ڣ�ս�ҵ�����ԭ�˿ڲ�����Ǩ���Ϸ��˿ڴ���ǰռȫ����20%��չ�����ڵ�Լ80%(������10)��3000��������仯���й��İ�ͼ�����Ʊ�Ǩ��ԽԶ�ţ��������滷�����������Ȼ������Ӱ���Խ����(1) |
")
{kind=link}
{kind=link}
{kind=link}
{kind=link}