![]()
|
|
|
�� |
�л�Դ����Ҫ <15>
<14> <13> <12> <11> <10> <��> <��> <��> <��> <��> <��> <��> <��> <��> ��ѧ���� Ī�粨 2008
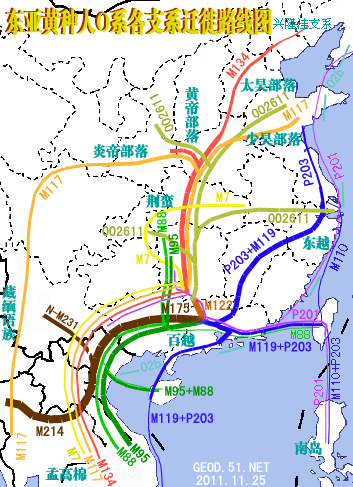
��������.��.�� ������ܺ��ѿ�����ʼ�ٶ����ĵĵ���Ⱥ����M134Ϊ���ģ�Ȼ����7ǧ��ǰ�ڻƺ����ηֳ���M133Ϊ���ĵIJ������弰��M117Ϊ���ĵĻ�����Ⱥ�����ں�����M117��Ƶ���������£��Ʋ��Ƽ���Ӧ��M117Ϊ���ģ��������ƺ����䵥����ӦΪM117����������˵�Y-SNP���ȴ¶���ڻƵۼ������棬���ܡ�������Ρ�Ӧ�յȾ���M122���෴������ΪԴ�Զ�����껺��������ȴ��M117��������յȡ��Ƶۼ��շ�֧���ܡ��⡢������Ρ�������¡��š��ơ������衢�Ρ�֣��κ�����������롢���������ڡ����������̡������ϡ�³������ë�������ӡ��ܡ���֡�Ҧ������ղ���ϡ����������š������ܡ��֡��ϡ��ܡ��Ρ�ʯ��Ӻ�ȣ��۽��շ�֧�н����롢�����͡������ꡢ�ա��ˡ��¡������ߡ������ء��ޡ�л�����������ġ��ޡ�¬���ˡ��������𡢶��ȣ��������շ�֧�лơ��졢�ԡ�����������ء�̷�����������������ӡ��𡢷ѡ���ȡ��ѡ��ĵȡ������Ϊʲô�� ��������.��.���� ����Ǩ�����ޣ�ʢ���ڸչ������������Լ��1.4����ǰͨ���������ǺͰ���˹�Ӱ뵺�Ĵ�½�ŵ��ﱱ�����������ո��ݵ�һ����Ѩ������һЩ�������㻯ʯ�Լ�����������������1.4����ǰ(����ѧ��)��������������ά˹С���ֵġ�����ά˹�Ļ���,����������1.33��1.29����ǰ(��ѧ��)������ĩ�μ���ڵ��ﱱ�Ǻ������綫����D��ʢ���ڵ���ʱ���DZ����������������Ϸ����Ӷ��Ʋ⣬������������Ӧ�Ǻ����ٴ��Ϸ��عź������ϵ��綫����C3������1.29����ǰ��һ��ֱ��Լ����ǧ�������ڱ����Ͽձ�ը�����������������ʯ���������˲������ͬʱ�������ٴγ����˳���1000��ġ�������(������)����ĩ�α��ڽ���(1.17����ǰ)ǰϦ��ӡ�ڰ��˵����ȵ����˱����������غ�������Ǩ�㣬���ܿ쵽������������˹ɽ��(��³����)��1����ǰ�ͳ�����ԭʼũҵ��ӡ�ڰ��˵�Y-SNP��ҪΪQ3���Ժ������綫����C3Ǩ�뱱��������Q��C�ֲ�������Y-SNPƵ����ͼ>>>��������N��O��1����ǰ����ʱ�����ں�ƽ�������ߣ���˿���δ�ܿ�Խ�����������ڱ��ǵ�Ǩ�㣬��һ����ֵ�ÿ��ǵģ����ǡ���������ͬγ�ȵ�����ȣ��ҹ����ϵ�������������ƫ��5�桫18�桱��(����Ǩ������ʱ������,����2�����ǰ,��ͼ>>>)(37) ��������.��.���� �綫����C3���������C�غ����ӷ���Ǩ����Լ6����ǰ�͵��ﶫ�����غ���C2��C4���˰��ޣ���Ϊ�������������������綫����C3��ʼ�ֲ������ϰ뵺�������弰�����������ȵص��غ���ӡ���һЩ������Ⱥ���нϸ�Ƶ�ʵ�C��Լ3����ǰ�����ڵ�������������ԽO2a-M95�����ϰ뵺���ż�ѹ���������а�ԽO1a-M119�Ի����غ��ļ�ѹ���綫����C3��2.5����ǰ�ͱ���Ǩ�㵽�����غ�����������������ʢ���ڵͺ�ƽ��ʱ����̨���Ϊ��ɽ��ij�Ա��C3��Լ��2����ǰ�͵����˶������������ּҸ�(22kaB.P)������ǰ����(19kaB.P)����������ɽ(17kaB.P)����ַ����������ʢ���������ڼ���������֤�ݡ�C3�ٲ��������ʰ뵺��ͬ�綫����D���ŵͺ�ƽ������ձ�����C3������Ǩ�㣬�������ٲ�����1.4����ǰ���ﱱ����1.29����ǰ�������Ͽ����DZ�ը������C3�������Լ1.8����ǰ��Q�����ǵ�������ֻ�����Q3������Ǩ�㵽���������Ƕ�������Լ1.2����ǰ��Q3��Խ���ޣ���Ϊ����ӡ�ڰ��������Ժ���C3β�浽�ﱱ����Q���ֻ���Q1���Ͻ�����ԭ�����Ϊ����ĵ�Ƶ�ײ㡣Լ1����ǰ������O2b�ض��������ϵ��ﶫ�������ʰ뵺������ԭ�������Ļ�����N���뱱�ǣ�����ԭ��ɽ�����ȶ���O3-M324Ǩ�㳯�ʰ뵺����ʱ�������ѳ�Ϊһ����C3��O2b��Q��R��N��O3*�������ɵ��綫���ˡ������ˡ�������Ѫͳ���ӻ⣬5��4ǧ��ǰ������O3e-M134��Ⱥ�����붫���ǡ��������ȵ�ԭʼ���ij�ԱӦ��C3�����ϲ�������O2b���Ժ����䱱�漰���������N�����Ǻ���������ı��ϼ����������Ϊ��������˸߱�����O3����Լ4ǧ��ǰ��λ�ں����������ε�һ����C3ռ���Ƶ���Ⱥ������C3ͻ���C3c��������������ͨ��˹������չ��ͨ��˹Ⱥ������������Ǩ�����ɹŸ�ԭ��չ���ɹ�Ⱥ�������ɹ�Ⱥ������������N��ӡŷ������R1�����ںϣ��γ�ͻ��Ⱥ�������������������ͼ�� ��������.��.��0 ̨��O1a*-M119ԭס��д��й�����O3*ΪO3��Դ�������ṩ����֤������ȵ������������������O1a*-M119Ϊ�����̨��ԭס���ɽ���Լ��2����Ӵ�½�ֳ���Ҳ����˵�������dz���ʢ���ڵͺ�ƽ�浽��̨��ģ�����ƽ�����������½��������ɽ�����൱��Ƶ��O1a*-M119���У��д��Ź�����O3*(��O3a5-M134��O3a4-M7)������������O3*ռ27.3%����������ռ14.3%��̨�����弰���ϸ�ռ9.1%����ЩO3*��O1a*-M119һ��������2����ǰ���ѵ���̨�塣���ڳ������ξ�����Ⱥ����Դ������IJ������������ڡ�������Ⱥ��ϵ�Ŵ��ṹ��̽��һ���������ᵽ:��5��Y-STR�ظ����ķ������M7���������������10300��11800��֮ǰ����һ���ƿ�Ϊ����ϵͳ����Ⱥ��ķֻ�ʱ���ṩ���ա�;��O3������Դ˵��������ȺO3*+O3a4��Լ��1.2����ǰ���س������£���������ɢ�������غ�ʱ��ƽ���������ߡ���ˣ��ɶ϶����ϴ��ڹ�����O3*������2����ǰ�ͻ�ͬO1a*-M119����̨�嵺������O3*��Դ�����㱱����������Դ�����ϵ�������֤�ݡ�(35) ��������.��.���� ������ӦΪ����Խ������ݻ��������˳��ֵ�����Ƶ��O3*�����ж�������Ϊ��Խԭס���������������֣��й�ѧ���Ʋ⣬�����˵����ȿ����Ǵ���ս��ʱ����ԭǨ�����Ƚ����ĺ��������缸���꣬��������Ϊ���ˣ��껰Ϊ������ġ��ʯ��������ɽ��ѧ����ѧϵ���������������Ĺ����������һ�Ŀ����������Ϸ������ս��Ĺ�ȣ������Խ��Ĺͬ����Խ�Ļ����ͣ�����̷Ԫ������ġ��-����,�����Ļ��Ŷ��ۡ�һ���323ҳ��ָ:���Ϸ������40��ս������������Ĺ����Ĺ������ƽṹ��������Ʒ���ص㣬δ���������Ե���ԭ�����ط�����������ͭ��������Ϲ�ϵ����������������һ�������Ĺ�Խ��ʹ�õ�������ٽ����ԡ��IJ��ȶԽ����˵�Y-SNP����������жϣ������˵�����ӦΪ����Խ(����+���g)�������������Խ����ԭ���ˡ���������ԭ������O3e-M134���֣���������ء����أ�Ӧ��O3d-M7���ҽ���Ӧ�������������׳�����ڽ����˳���̹�˵�����ǵ�O3ϵ������O3d�н�Ե��ϵ������Խ��O3ϵͳO3d-M7��������Խ�����ټ���̨��Ⱥ��O1a-M119�� ��������.��.���� �һ������O3ϵ��������Ⱥ��O3�ĵ��������ģ���ηֻ�Ǩ�㣿̤����Ь���ٴ���û�뵽���-����һ���Ľ����˾���O3*������Ⱥ���й�Y-SNP������Ϊ��O3��3����ǰ���ѵ�����������O3d������O3e�ȵij�ʼ������Ҫ���ö࣬���ܲ�����1.5����ǰ����ˣ��м��Ӧ����һЩ3��1.5����ǰ���µ�O3*������Ⱥ���ҵ���Щ������Ⱥ�����п����ҵ�O3�ĵ����ء����ǡ�������O3*�ľ���������������ΪO3e��O3d��O3c�ȵ�Ⱥ���ɷ֣�����O3*��Ƶ��Ⱥ���������ϲ����������弰�ϵ����������������Ƶ��O3*�п���Դ����ս�����ӵľ�����Ⱥ����O3d��Ⱥ���ɷ֣����ϵ�������O3�ĵ����أ������������൱������O3*��������O3e��O3d������һ�����Էֱ��Ƿ��ɺ��˻��������롣�����˵�������Ƶ��O3*ΪO3������Զ�ŵķ��g�����ṩ��������Ⱥ��������ݡ�������ԡ��IJ���������������ģ����컳�������˹���34��������O3*�ߴ�52.3%����O2a*ռ17.7%��O1a*ռ14.7%��O*ռ5.9%��Cռ2.9%��������O3e����O3d��O3*������Ƶ�����й����Ժ�����о����������ž����о�Ա�����ġ��껰�о���һ�飬�껰�Ǻ�����ϵ��̨���嶱ˮ��֧��һ�����ԣ������Ǻ�����κ�һ�����ԣ����Ƕ�̨����һ�ֶ��������ԣ����ҹ��·��������еĻ��ʯ(-�����»�����.2003)���㶫ʡ�����о�������־�о�Ա�ڡ��㶫������γɼ���������׳�������ںϹ�ϵ����ָ��:�����������ԡ��껰��,����ѧ����һ�¿϶�����׳�������һ�����ԡ�������������������Թ�ϵ��ʮ�����У���֧������δȷ�����ر�ֵ��ע�����,���껰��Լ��1/3�ʻ���䱾�������У�Ϊ׳��������������������������Ȼ�����̳�׳������������Լ���̨��ײ�����Է�յĻ����ж�����չ�Ľ�������������������,���껰�����ŵ�ǰ�����Թ�������һֱ��������һ���Ĺ�Խ��ĺ��ᡣ����������Ҫ������-��ɽɽ���Ķ��༰���࣬�ֲ��ڻ����ص�ʫ��-����-��ͷ-���-�����Լ���صij���һ�ߣ����ݼ���ɽ�����У����˿�Լ15�����л���Լռ10������˼���ǣ��ؽ����εĺ��ݡ�����һ�����ײ�Ԫ�ر������ĸߣ�������15������������O*��60%������28��������D1��18%��(��2009�꡶�ִ�����ѧͨѶ�������ڳ����¡�������Ⱥ������Nϵ����Ⱥ��Ǩ��ֻ���һ�ģ����������˼�������������Ϊĸϵ������A����Ⱥԭʼ��A*�ֲ�Ƶ�ʺ��ڲ�����Ƶ�ʵ��ص��߷������������ڲ�����Ƶ����ߣ���8.8%����������Ϊ7.7%����һ������Ϊ��A����Ⱥ������ֻ��أ���A�ķֻ������Լ��4����ǰ���ɼ�������ӦΪ������Զ�Ź�Խ��ĺ��ᣬͬʱҲ֤��O3�ĵ������ںؽ������ε���) ��������.��.���� �����й���ԭסO3-M122������ԭס��ҪΪO1a-M119��O2a-M95�������ϵ�O3ϵ�����ɱ������ĺ��˼�����-���ϵ���������Ⱥ������ڴ��룬���������M122/M134��ֵ�����1�������ϡ�ɽ���ȵ����ھ���-����ԭס��M7��M159�����������ҪΪ����M122�������뺺�����M122/M134ƫ�ߣ���ֵ��1��1.5֮�䡣��������(�����ӵ�����)��ԭסO3-M122��M122/M134��ֵ�п��ܳ���1�����о��������ĵ������ܻᳬ��1.5�� ��������.��.���� 2.5����ǰĩ��ʢ���ڵĵ��������㱱���ѷֻ���Oϵ��������Ǩ�㣺�µ�������ʾ��O2��O1��O3�Ⱥ�ֻ�������ܽӽ�������Ǩ�������ΪO2a-M95����Ҫ�ӹ������������ϰ뵺����Ǩ�㣻���ΪO1a-M119����Ҫ�Ӷ����غ�����Ǩ�㣻O3ϵ��Ǩ��ģ��С��������Ҫ����O2a��Ⱥ֮�У����Ա��ҪΪM122��ϡ�ٵ�M121��M164��ʱ����Ҳ�ѵ������������У���M7��M134��Ǩ�������Ӧ����1.5����ǰ�����ǵ�����O*��Ҫ������O2a��������¡�˵�������ʾ��O1a��ȺҲ����O*��M119��Ƶ���������������塢���塢ˮ�塢�����塢�����塢��ɽ�塢׳�塢����ȡ�������¡�˼�������Ⱥ��M122��Ҫ��M119���(����>>>����>>>����>>>)������˵�����ڵ�O3�����O1a�Ӷ�������Ǩ�㣬������Բ�ʿ��Ϊ��Խԭס�ʯ�ĺ�������ȴ��δ������O3*����:����ʷǰ�Ļ���ַ����֧Ǩ��·��ͼ������OϵǨ��·����Ӧ���ʾ��ͼ(31) ��������.��.���� ���,��ȵ��ڹ��ʶ��������ģ����ϰ�ԽO1a-M119����ԽO2a-M95�Ĺ�������ֱ�Ϊ3.6���꼰3.2���꣬��ԽO2a�Ĺ������仹���ܴ�4���ꣻ��ԽO1a��������Ⱥλ�ڹ㶫������Լ2.2����ǰ�ֳ�̨��֧������֧��O1a����֧��ֱ�Ӵӹ㶫����İ�Խ��Ⱥ�зֳ����˽����ʹ��ԽO1a�����������ԭ�����Ƶ�2����ǰ��Ϊ��ǰ��[��ʱ�估�ص��ϣ����ӽ����ɳ�ҵڶ��Ļ���(48��5kaB.P)����۴��;�ʯ�����쳡(35��39kaB.P)]��ͨ���Ժ�������O2a*��O1a*�ķ����ó����ۣ�ԭʼO2a*��Ⱥ��Լ��2.6����ǰ�����ڵ������ֵͺ�ƽ��ʱǨ�뺣�ϣ�������O1a*��Ⱥ��̨��֧�ֿ����Լ��1.8����ǰ���뺣�ϣ�֮����ƽ��������������Ⱥ���½���룬����ʯ��ʱ��������������ԽO2a����ԽO1a��Ⱥ�ɺ����뺣�ϡ����������O3�൱ϡ�٣��ҿ��������ڻ��룬˵�������ܱߴ�½��û��O3������>>>ԭ����ԭ������ͼƬ����(30) ��������.��.���� �ܴ�����ĸϵGMѪ�ͷֲ�ͼ������ʲô��������ͼҲ�������˴���۾����������ţ�������Ⱥĸϵ���г��������綫����Ѫͳ��������Ҫ����С���ڡ��侲�����룬������Ҳ����û�С����˵�Ǩ���������Ϊ�������ҳ�������ս����ʤ�����ץ���Ů�˵�ս��Ʒ������ѵжԵ�������������𣬽����ԭסĸϵ�Ļ������ر�����������������ϵ�����������������棬Ҳ�ɴˣ�ĸϵ������ܷ�ӳԭ����������ò����ͼ���ɿ�����ĸϵ�Ϸ��Ի�����ռ�������ƣ��������綫���ˡ������˸�ռ�൱��������ϸ�ϵ��������Ϳɵõ�һ��������Ǩ����̣�Լ5����ǰ������Գ���Ϊ�磬������ռ���Ϸ������綫����ռ�ݱ�����1.2����ǰĩ�α��ڽ�����������Ϊ������Ϸ������˽�����������������������˱������������ԣ������˴��Ů�ԣ�������ϵ������ȡ���˾������ƣ���ĸϵ����ԭ�ȵ��綫����ѪͳΪ���������غ�ʱ�ڣ��������˴����ٽ������ϣ�����������˲��ְ�Խ���ԣ�����ԽŮ�������ر��������������������ɰ�Խ����Ϊ�Ժ���Ϊ������ĸϵ����ԽѪͳռ���ơ��۹ۣ���ϵѪͳ�������������ɱ����ϣ����˸����أ������Ļ������滻��ԭס�綫���ˣ����Ϸ������˻��ǻ��֣�ֻ�Ǻ����滻�˰�Խ��ĸϵѪͳ�������綫����Ѫͳ�������ɣ�ֻ�Ƕ�����˻��֣����Ϸ�������Ѫͳ���ǻ��֣���Խ���ǰ�Խ�� ��15�� ��14�� ��13�� ��12�� ��11�� ��10�� ������ ������ ������ ������ ������ ������ ������ ������ ������ |
{kind=link}