![]()
|
|
|
�� |
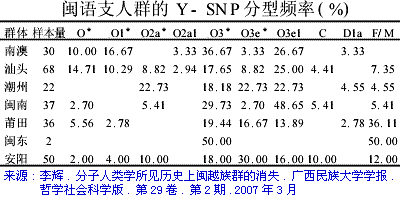
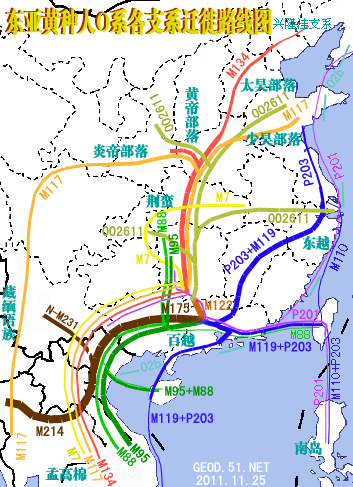
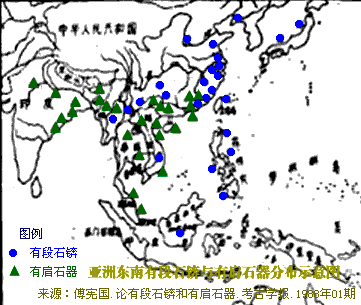
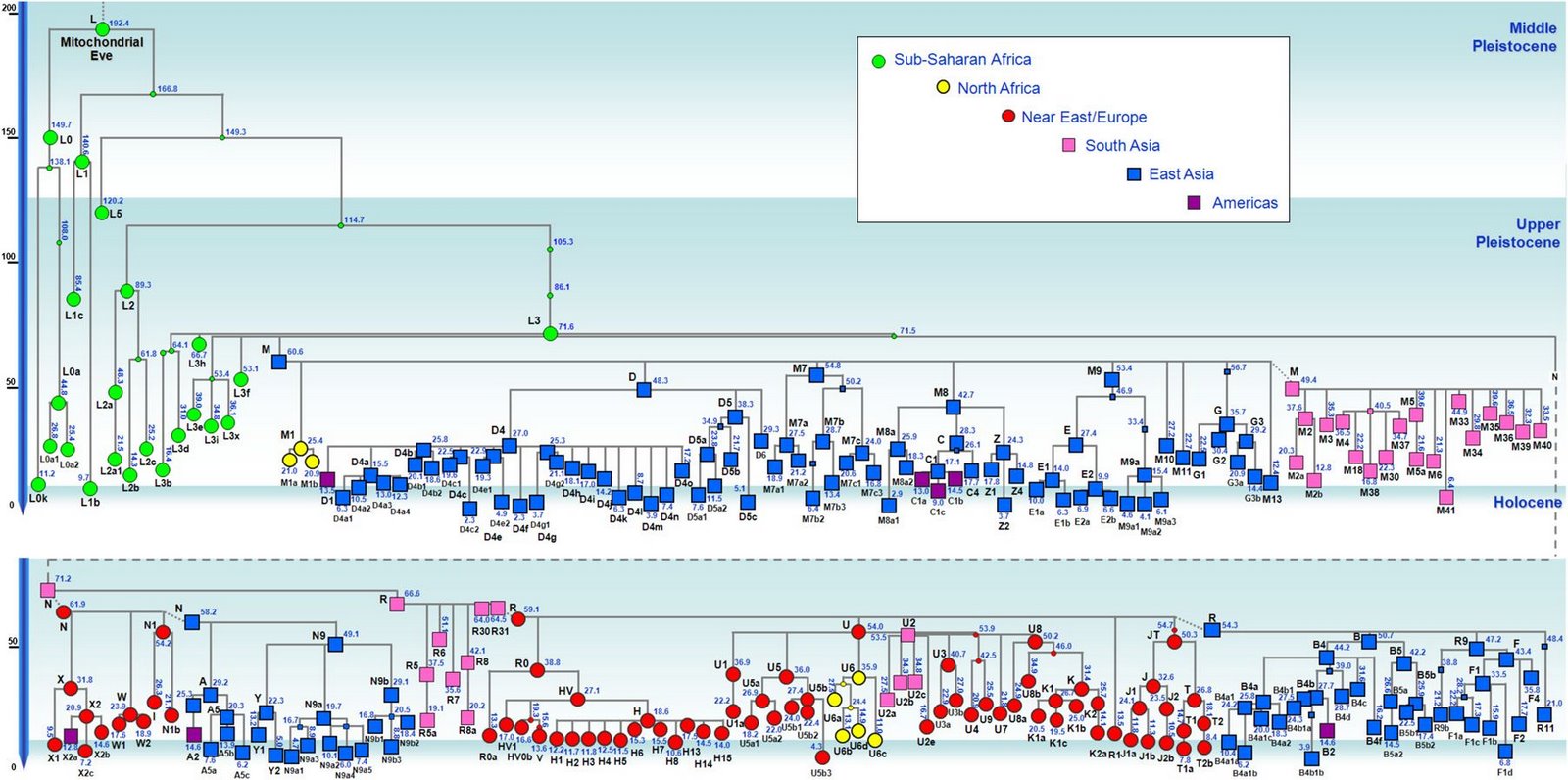
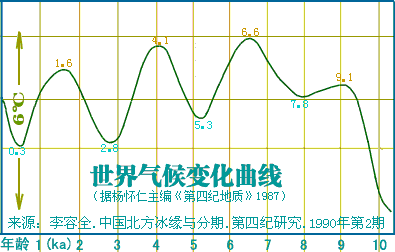
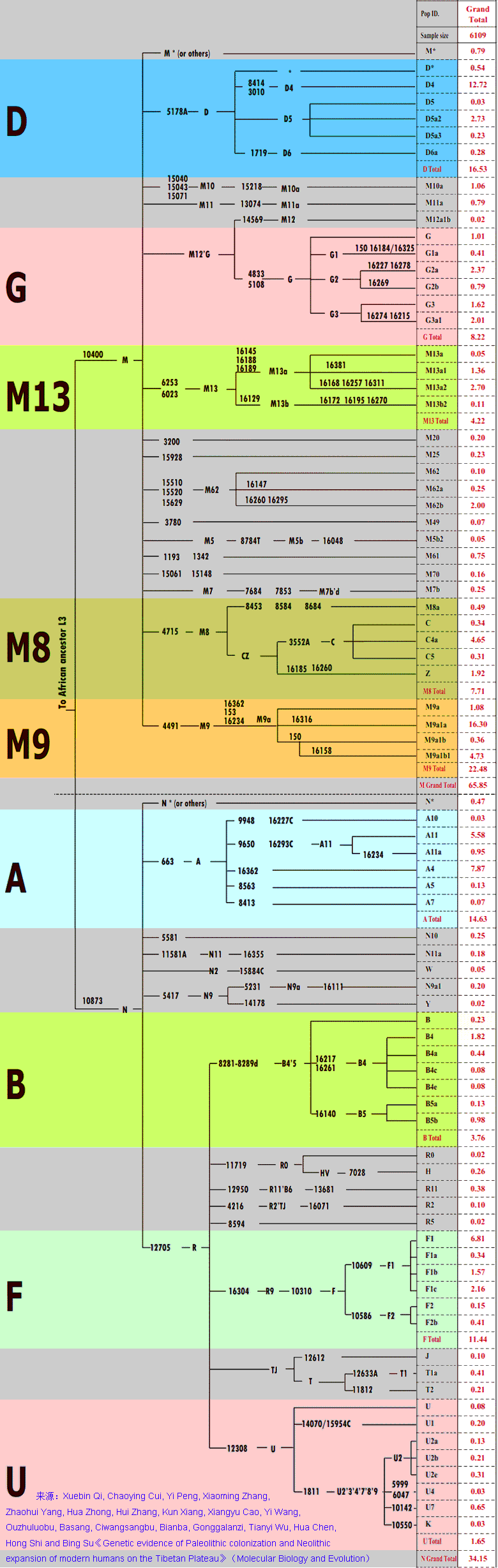
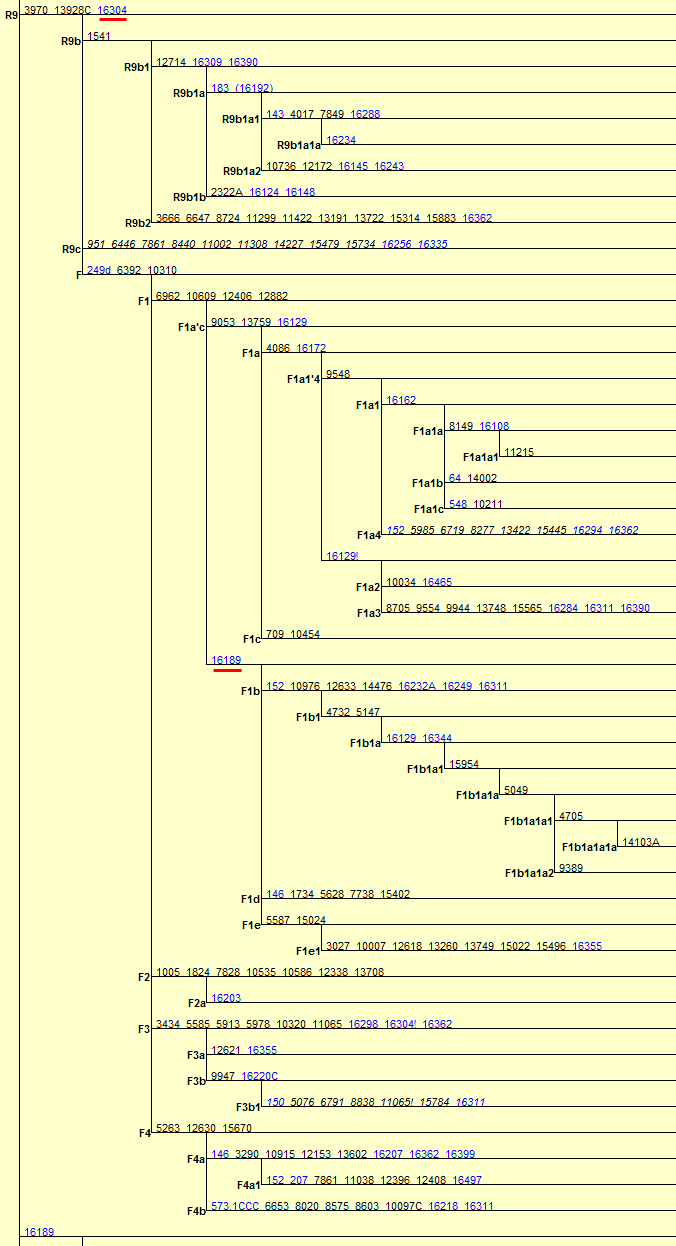
�л�Դ����Ҫ <15> <14> <13> <12> <11> <10> <��> <��> <��> <��> <��> <��> <��> <��> <��> ��ѧ���� Ī�粨 2013��2014 ��������.����.���� ��002611��Ƶ�ʷֲ�������Ⱥ��ϵ��002611��Ϊ���������Ƶ�ĸ�ϵ����Ⱥ�������ڳ������ϵ�ԭס��Ⱥ���൱��Ƶ��������ʵ���������������������Ϸ�����׳��21������������27����������11����һ��ֻ����1����ռ1.7%������60�������51����������23����������11����һ��145��δ��һ��002611�����Ϸ����嵱�У��㶫����40������������59����ƽ����101������������13����һ��Ҳֻ��7����ռ3.3%�����ϰ뵺������1133��������18����ռ1.6%����002611��������ռ20.7%(12/58)����Խ������ռ14.3%(10/70)��̩����ռ11.8%(2/17��������)����������ԭ�����ϣ������ԭ�ӳ������ε����(λ��ƫ��)������ԭ�ӳ�������λ��ƫ����������������Ҫ�����ϰ뵺���ϸ���(�ݲ���������)����������002611ԭ���ڳ������Ϻܿ�����Ƿȱ�ģ����е����DZ��������Ժ����ı����������002611�п������Զ��ĵ�̫껼��ţ����������϶������Ƽ��Ŵ�ս�����ĺܿ�����002611��һ�������������������002611Ӧ�Ƕ��ļ�����˫��Դ�ģ��������������а˾�ǧ�ꡣ��ɽ�Ļ��ڣ���껼��ű��Ƽ��Ŵ�ܲ��沢����������Ң˴��ʱ�ڻ��DZ����������Ķ���(˴����������Σ)����ˣ������002611��̫�����ǻƵ�������Ի��ļ��ţ���������û��002611������Ҳ�ͳɲ��˱������������002611�ƺ�ֻ�������ڶ���(ͬʱ�����M7Ҳ��������)����ص���ԽӦ������������ȣ���δ���붫��������Ƶ�ս�������ں��������ͬ�����γɳ�[ע]������Ӧ�Ǻ����˼��ձ���002611��Դͷ��Խ���˼�̩���˵�002611�ܿ������غ���������ĺ������������ԭ�˼����ˡ����ۣ����������ձ�����������Ⱥ(������)��M117Ӧ��Դ�ڶ��ĵ���껼��ţ����ڲ����O3ϵͳ��ҪΪ�������M117(29.82%)����M134*��Ƶ(2.63%)(����ѧ��)����ˣ�ǰ�����Ļ��ڣ�M134*�ı�����Ӧ���ڹ��к���������ǰ�������ų����Ʋ⣬���������Ƶ�ĸ�ϵ����ȺM134*(F444)������Ӧ�����ںӱ��Ĵ�ɽ�Ļ������ڵĺ���Ļ���Ⱥ��O����Ⱥ��ϵͼ>>>��O����ϵ�����ֲ�ͼ>>>��(109) ��������.����.�� ���ڶ�����Ⱥ��M117��M134*����Դ���Ϸ���������Դ�Թ㶫������������������һ������ϵ����Ⱥ��O1a-M119(P203��M110)Ϊ����İ�Խ����������ʯ�������м�ʯ�����ж�ʯUĴ����жϣ��Ļ�Դͷ��Ҫ��������һ����Խ����ɽ�Ļ������Ƕ�Խ��ķ��-����Ļ�����Խ��������̨���M110����C3*������Խϸߣ�����Խһ�㲻�������ֵ���Ⱥ����Լ��1.5����ǰ������M119(P203)��Ⱥ��һ֧Ǩ�������㽭���γɶ�Խ�������ӹ㶫��һ֧�������Ϻ�����ɽһ����÷�չ����Խ����Խ�ķ��������Լ��1.5����ǰ����Լ������ǧ��ǰ����Խ����Խ�����ܱ߲����˶���������Խ��Ҫ����Ϊ�ж�ʯ����������γ��ϵ����壬��Խ��Ҫ������˫��ʯ�����ܱߴ�����ʹ��Խ�������϶�������������ʯ�������ڹ�����û��M119�ģ���ˣ�����û��M110��C3*�Ķ��������Դ�Թ㶫�������ٸ���Ҳ�ǣ�������M110��C3*�����ϸߵĶ���������Դ�Զ�Խ�� ��������.��.���� �����Ͽ��������ڹ�һ�����Է�����ǰ�������ˣ�����ѧ�������ϵ���������Լ20����ǰ���ڷ����֣�����ѧ�ϵ��ִ������ȴ�Լ��6����ǰ�߳����ޡ���ȫ����ɢ�������й��Ϸ������ֶദ7��13����ǰ����ǰ���������ż������Q��������ϵ����������˻�ʯ�Ϸ��ư������Ϊ7��13����ǰ����磻�����������˶����ǻ�ʯ�����������������������˹��ɽΣ������11����ǰ�������������ɺ����˽������ⷨ�ⶨ��������7��15���ꣻ�������ڹ�����ɫ�ﶫ½�Ƕ��ھ����������˳ݻ�ʯ�������7��12.7����ǰ����������5��10����ǰ���ż��У����ɳ����ַ�����Ļ���(��ϵ��79��15ka)�����ⷨ7����ǰ����Ͽ��ˮ����ַ(�����ĵ�)����ϵ��4.7��10.5����ǰ�������������˶���ַ(���·��)����ϵ��9.4����ǰ��������������ַ�ȡ�������ǰ����������7.4����ǰ����������������ˣ�������δ֪�Ļ������Ŵ����ִ��ˣ����ڲ������и�Ƶ(90%)�ĵ��������˻���EPAS1(������������)����ˣ���������-�������ִ������ȵ���ʱӦδ��������븸ϵD��Ⱥ�������ںϣ��ɴ��Ʋ⣬��γ�Ϸ�����ǰ�������˸�û�����������ϵΪD����Ⱥ�������ں�����ǰ�������˵Ļ���ֻ�����ֻ�����Ŀǰ��δ��ʶ��ͬʱ���������е�Ƶ��EPAS1�����ų�Ҳ�Ŵ��нϸ߱�������ǰ�������˻��������������ִ�������û���뵤�˷����Ӵ���ֻ�ǵ�������ǰ�������˷������ӽ��������е�Ƶ��EPAS1�������ִ�����������ǰ�������˵��ںϡ�Ҳ����һ��������ġ���Խʱ�յ��ںϣ��������й����ʯ����һϵ����̬�ϵĹ������Ƿ�����Դ�뱾���ӽ��Ľ����Ŀǰ����������ѧ˵��һ�㣬������������Ⱥ�ĸ�ϵ��ĸϵ��Я���з������ȵ�ͻ�䣬Ȼ������仰Ҳ���������������������е��˶��ں������Է������ȵ�ͻ�䣬�����ų���������� ��������.��.���� �����ļ��о���2014���1�ڿ���ר���еĹ�������Ϣ ��.�����ǡ����������ȫ����11��8kaB.P.��ĩ��ʢ����֮��ĵ�һ����ůʱ�ڣ�ȫ��ƽ������Լ60m�����ڸ�ʱ�ڵ�8.2kaB.P.������һ�����¼���ʯ��ׯ������ʱΪ��Ϊ��ʪ����Ҷ�ֲ�ԭ��������.���ԡ�Ī���ŵ���ͨ�����±����ȫ����������ܽ�ͶԱȣ���������仯������Է�Ϊ3���Σ�11.0��8.5cal.kaB.P.���ȶ��������ڡ�8.5��3.8cal.kaB.P.��Ϊ�ȶ��ĸ������Լ�3.8��2.2cal.kaB.P.�����ڡ�4.0cal.kaB.P.ǰ�õ����Ĺ��Ļ��쳣���٣��������ֵ�������ַ�������739�������������������400��104�O��ʯṹųǣ���֮����������ǧ��֮�õ���ͭʱ������ַ����ȴ������٣����̺�����ʱ����ַ���ֱ����77����52������.���ա���һ��������ʵ�������Ҥ�Ļ�������ʱ�ڣ�����Ҥʱ��(5300��4800cal.aB.P.)��ַ������530������ɽʱ��(4500��4300cal.aB.P.)��ַ����(249��)�������٣���������ʱ��(4300��4000cal.aB.P.)��ַ����(674��)���������ӣ��ҷֲ�������������չ����ɽʱ�����������ʱ�ڽ�ˮ�������ࡣ��.��骡���Сǿ����������ˮ����5050��4870cal.aB.P.����ů�´������ȴ����ɵ�������ϣ������ԼΪ12.9�棬�����ڸ�Լ1.5�棬�����ˮ��ԼΪ758.4mm�������ڸ�Լ180mm����.���ա�����÷����ɽ����ɽ�Ļ�����(4500aB.P.��)���ܴ��������Ľ��¼�����ͻ���¼�����ɽ�Ļ�����(��4200aB.P.)�Ʋ��������ůʪ����ɽ�Ļ�֮�����������䡣��.������ɰ����һЩ���ȴ����ֵij�����ʾ¤������Ļ�ʱ��(4300��3800a.B.P.)��������ܱ�����ůʪ�� ��������.����.���� ����ԯ������ͬ�Ĺ۵㣺��.½Ρ���ⱦ³1997���ڡ�����ѧ�����Ϸ�������������ʯ���������ȹ��ɵ�����ϵ������Ϊ�����Ƥ��-����ͷ�Ļ�����ԭ�Ļ����и��������ϵ����������������Ϲ�̨�Ļ���Ϊ���ơ��ɴ���֪���Ϲ�̨������Ǩ����ŵأ�������Ļ����Ƥ���Ļ�������Ⱥ�����죬������Ļ��ɱ��Ƶı���ͷ�˺Ͳ����Ƥ���������죻������Ļ��DZ���ͷ�Ļ��ļ̳��ߣ������Ƥ���˽������ǰ���ں���ͣ���ӣ�����������Ļ��Ĵ��죻ͬʱ������������ϵ��˵������ͷɽ�Ļ����-���Ļ����Ƶ�һ����ת�㡣�������ڰ��¹�����ѧ֤��(ͷ����̬��)Ҳ˵�������о��������ǵ������Դ��ڶ������ˡ���.����ҵ2009���ڡ���ԭ���š��Ϸ�����������Ϊ�ֺ��Ļ���������Ļ���ʵ�Ǵ�ͬС�죬������Ļ�������ò�����ڼֺ����ڣ�������Ļ����������μ������ͺ�ˮ���Ρ�����Լ���ԥ����Ӱ������൱���ԣ����Ļ�(�Ϲ�̨�Ļ�)������������Ļ�������չ���������Ļ��ںϵIJ�����첿�֣�����Ļ������ơ���ֲʡ�������ѹ�⡢����ѹӡ���ߵ�����������Ͽ������ͥ�������DZ�Ϫ�Ļ����ƣ���ˮ������������Ļ�������ͨ����������Ļ����ڶԴ�ɽ�Ļ���Ӱ���������ԡ���᷵���ԭ���ֲ��ź�������Ļ�����ͬʱ�ĺ����Ļ���Լ��Ԫǰ5400��ת��Ϊ�����Ļ����������еı����Ļ��Ŵ涼�����ٰ�����������Ļ����أ���������������Ļ�������Ⱥ�Ķ���Ǩ�㣬�Ŵٳ��˺����Ļ������Ļ�ת�䡣��.�ų�2011���ڡ�����Ϸ�����������Ϊ�ֺ�һ��ƫ��ʱ����Ҫ�ֲ��ڻ��Ӻͺ�ˮ���Σ�ƫ��ʱ�����������ֲ�����ԥ�к�ԥ�����Ѹ�����������֪��������Ļ��ֲ���Χ��������Ļ�������ȺӦ�����Լֺ�һ�ڣ��ֺ�һ����������Ļ�����ҪԴͷ���Ϲ�̨�Ļ�(����Ļ�)����������൱��������Ļ�ƫ��ʱ�ڣ��������͵������ӣ����ڹ��е��������Ҳ���Դͷ����˺ܿ����ǰ�����͵ĺ����Ļ������������Χ������ɽ�����������Ļ���ʱ����������Ļ��൱������ȺҲ���Ը�������Ϊ����ʳ���Ŵ���Ҳͬ���е���ֲ�����Ҳ������ֺ�һ���Ŵ�ͬһʱ�����Ƶ��Ϸ��Ļ�ϵͳ������������ƺ������ε���6000BC�Խ���ʯ�������Ļ����γ���Ҫ�dz�������ͻ���һ���Ļ�����Ⱥ�����Ľ����>>>(105) ��������.����.���� �ǿ��ˡ����ˡ��������ִ��� 12��4�ա���Ȼ����־�����Ŵ�ѧ�����������������ǿӡ���ַ��һ��40����ǰ�������ȹ��У���ȡ����������ϵĽ�����������������������У����������ʾ�������ᰲ�����˻��ִ����࣬����DNA�뵤�������˸�����������뵤���ڴ�Լ70����ǰ���й�ͬ�����ȣ�����ͼ���˽����ʾ�����������˿��������ж��������롰�ǿӡ��˽�Ե�����ڶ���78����ǰ����ײ������֮���߳�������Ⱥ�����>>>������ڡ��Ŵ�ѧ���˿���ϵ���ع�����ָ������������ϵ�ִ��������˷ֿ�40�����꣬�����뵤�˷ֿ���Լ100���ꣻ�ڷ���֮����ִ���Ⱥ�У���������1��4%�����˻���ɷ���������������Լ7����ǰ���߳�����ʱ�������û�з�������������Ȼ�����ڱ���(����̩ɽ��)����������½�ִ���Ⱥû�з����κε��˵��Ŵ��ɷ֣������ڴ������¼��������������˴�Լ6%���Ŵ����������������ǵ�������Ǩ��;�����ϰ뵺ʱ�Ӵ����˵��ˣ��ƶ��������������뵤����ͬһ�����֡�(104) ��������.����.���� ��̽-�����Ļ���������Խ�Ļ��ĺ���������Խ�ǿ����ĵײ��������Ļ�����������ǧ�����⣬�������ܱ��Ļ�����װˮʯʯɽ�����˾��Լ1����ǰ��ʯ����ַ(��̷Ԫ��)����Ӧ��Զ�Ų���Խ����������������Խ�dz�����ʫ��һ�������˵�����(��������)��Ҳ�ǿ���������ԭʼ�����������˸�ϵDNA������O3*��û���ֱ�����Ⱥ�����M134���������M7(�����)��ĸϵԭʼ��A*�ֲ�Ƶ�ʼ��ڲ�����Ƶ�����(��������)���Ʋ⽲��������ԭʼO3�����ֱϵ���ᣬ��������������O3�ĵ����ء��Ϸ�������ھ��38��ս��-������������Ĺ��(����ʽͦ)�����б��ڰ�Խ�Ķ����ᣬ��������Ĺ��ѧ���������Ϊ��Խ���ͣ��������Խ��Ǩ�йأ�ͬʱ��������Ĺ����ս�����������ھ���������˵������ʼ��ͳһ���ϲ�δ����ʱ�Ŀ�������ɴ��Ӱ�죬��건�սӦ���ںؽ����ؽ�����Ҳ������걾ӵء���Խ�ĵ���ռ�����������������Խ���ֱ���������-��ɽɽ���Ķ����Լ������ij�������ɽ�ȵأ����ǵĺ���������ڵĽ����ˣ����²��ֱ�ͬ�����������뽲�����кܶ���ͬ�����ף���������������Դ����Խ�Ĺų���ײ��⣬��������һЩ��껰��ͬ�Ķ�ˮ��(������)�ײ㡣������ɽ�����������������ƽ����Խ����Ԫ˧Ī���棬�����DZ��������ƾ������������ŵأ���¥������Ī���ڶ���-�ϱ���Ǩ��(������)��Ī��Ӧ�����簲�ӿ�������Ⱥ֮һ�������ƽ����Խ����Ī����ȱ������˴�������ԭ���ԣ�������ų���Ļ���ټ��϶�ˮ��ײ㣬�������ɴ��γɣ������������Ŷ����ɻ�ͨ���ܿ����γ��ڷ�����ʱ��(ǰ106��426)����Խ���������Ļ����������ڿ����ǿ����Ļ��������ܱߵĸ���ԭ����������������ǿ������δ�����Ǩ����Ⱥ����ͬ����>>> (103) ��������.����.���� ���������ڽ���̨Ͻ�����������8.2kaB.P��mt-E��mt-E���ϵ��������е�ĸϵ�����ͣ�����̨���ɽ�弰�ϵ���Ⱥ����½��Ⱥ���С���������C14�������ڴ�������ɺ������̨����п��Ļ�(��ǧ����ǰ)��ҲԶ�����ϵ���ʯ���Ļ�(��ǧ����ǰ)���ɴ˼������Զ϶���̨��ԭס���������Ļ��ϻ���ѪԵ�Ͼ�Դ�Զ����غ���ʯ�����������ڡ�ʢ�����������۵������ۣ�����ǵ߸��Եġ�E��M9������M9a���еķ�֧��������27.4kaB.P�����ڴ��п��Ļ��ķ�֧��E1-14ka��E1a-10ka��E2-9.9ka������൱����E2a-6.9ka��E2b-6.6ka��E1b-6.3ka�������½��Դͷ����Щ����Ⱥ�ڶɺ�ǰӦ������ɢ������½ֻ�м������ǵ�E1*������֤����������������mt-E����Դ�м��ֿ��ܣ�һ��E��Դͷ���ϵ���Լ����ǰ�ӷ��ɱ�Ư������������E��Դͷ��̨�壬�˾�ǧ��ǰ�ɺ����������ĸ�ϵ��M110���������㽭�غ�������Դ��ʢ����̨���̨�庣Ͽ�ľ�ʯ�������������Ǻ�ƽ���������ڹµ������д�������������˵����ޱʵ�Զ����Ư������ȥ���������ֿ����Ժ�С�������Ư���ɹ��ʸ��ͣ�����ѧ֤�ݱ����ɺ������������ǧ��ǰ���ۺ�(64)��(66)��(67)��Ϊ���ϵ�����ĸϵ��E�����븸ϵM110���Ƶ�Ǩ��·����E����������̨��ʢ��������(��������)��һ�����븸ϵM110�ں�ƽ������(14kaB.P)ǰǨ�㸣�����㽭�غ�����Խ����M110��Լ��ǧ��ǰ����ƽ���������������е����½���ϣ��������ڹµ������������������ų�����ʯ������Ϊ��Խ����Eû�ܳ����ڴ�½��Ⱥ����(������)�������˵���֫������Ҳ����������������ԽԶ����ԨԴ���б��ڶ�Խ����Խ�Ķ����ἰ����ϰ�ס�������������E�ķ��֣���һ��˵��̨�����Ļ�������ѪԵ�����ϵ������Դͷ��(102) ��������.��.���� ������DNA����ȺF���Ҳ�õ�F1b'd'e�����IJ������У�F*�����ϸߵ���Ⱥ�����ǣ�̨��̩����8/20��������3/10����ũ��9/46������������9/85���㽭����3/30��Խ����3/35������������3/37��������8/97������F*������������16.2%(43)����������10/100��������8.7%(103)���人��7.2%(42)��������7.1%(69)��̫ԭ��6.9%(58)������6109����������11.4%��F��Ҫ��F1*(ռ6.8%)����ϸ���������F*���й����ϼ�������1.4��2.4%����ӡ��������ޡ�Lonbok�����嵺��1.5��2.5%��������������F*ϡ�٣���Ҫ��F1a��F������Ϊ43.4ka��F1Ϊ33.5ka��F1aΪ22.2ka����Ƶ�ʡ������������F���ܵ����ڱ�����һ����һ֧��ON-M214�������ϣ�������Mϵ��ѹ��֧�����飬��ʣ�����ǵײ㣻��һ֧��Oϵ���齭ˮϵ��Ǩ�۾����ϡ�2.2����ǰ��F1a����ڹ�����O2a��Ⱥ�е��������Dz�����O2aǨ�����ϰ뵺���ϵ����γɸ�Ƶ��(��Ƶ������)����һ������㶫��O1aǨ̨�嵺�������غ�������һ������O3�������볤�����Σ�Ȼ�������ϼ�����Ǩ�㣬������һЩ��A��B��M7bcһ��y-D�ٳ���ʢ���ڵ����ձ�(�ض�������)�����ڸ�����ѧʵ���Ҳ�õ�������DNA��F1b'd'e(�����w��׃�^I�ĽY��Ϊ16042A��16051G��16183C��16189C��16269G��16300G��16304C����10310Aͻ׃)����16189��λ��δ֪�ġ�F1b,d,e����÷�֧������F1*����F1b������F1a��F1c��Զ��F1b������1.6����ǰ���й�������Ⱥƽ����2��4%���������������Ըߣ���Ϊ�������弰���壬��½������Ⱥ�ټ���δ����̨��ԭס������������������̩�����������ϵ�����û������ʾ��F1b�������Դ������O3Ǩ��(��A����)��(101) ��������.��.�� ��y-N���ɻ���y-N�Ƕ����֣���ĸϵ����ȺC��D4��G��M8a��M9a��M10�ȴ�y-D��Ⱥ���ﴫ����������Ⱥ(������������)����N��NO-M214�����η�֧�����ǵ�ԭ����ʲô��y-NOǨ������������·�߿��ߣ�һ���Ǵ�ӡ�ȶ�������������ϣ���һ�����ڱ�������O���ֺ��غ����ᡣ����������ᣬ�������ڵ�mt-N��δ�����ηֻ���·������y-D�������������Ⱥ��������y-D��Ⱥ��ֻ��1222����������F-M89*(xJ,H,L,P,O)��Ƶ(4.1%)(������)��NO-M214�ټ���Ѱ������ӱ�����Ǩ�룬���ڱ�������mt-B��F��N9��R9��M7�ȵij�ʼ���ŵأ�y-NO��ԭ���������Ⱥ���ɣ������F*��Ƶ������ζ�ź��������Ϸ�������ȫ�ĸ��棬y-NO��ԭ�䱻y-D��MϵŮ����ȫ�滻����һ���ɻ���y-N����ǰ��Լ����������������£�NO-M214ȴ�����Ų���ɢ�����Ǵ����������������������ģ������NO-M214(xO,N)����ںδ����ֵ�Ҫ�����ƺ�Ԥʾ��ʳ����Ĵ��ڣ��ѵ��������ϲ����������������ʳ�˷�ʢ�С�,�Ӷ����µ����������˿��������仺������أ�ʯ��ȷ�����������һ��ȷ��y-N���������������Ǩ��·����(100) ��������.��.���� ��������yD������Oϵ��ԭ�䣺������Ⱥ������Ǩ�����ںϣ��ټ���y-Dֻʣ�²��弰�ձ����нϸ߱��������ԣ�ֻ�ܸ�����Ӧ��ĸϵ����Ⱥ�����������һЩ��DNA������ԭס��������Щ�жϡ�y-D��ӡ�ȶ���Ǩ��ض������ϴ�Լ��5.5��4.5����ǰ����Ӧ��ĸϵ��M*��D��M8��M9��G���������Ƕ�δ�����ηֻ����������ֵҹ�DNA���������������о�δ���ֳ�A�����Nϵ��Ա�������Ϊ��y-Dϵ��ԭ��������һЩ���ϵ�Nϵ���ͣ���������Ҳû�������������������˵�pre-B���ձ��ض���������ԭ��Mϵ�⣬����Nϵ��A��B��F�Լ�M7b��M7c���Ϸ���Ⱥ�������ͣ�����Դ�ж��ֿ��ܣ�һ����5��2����ǰ������һ����y-NO��Ⱥ������ĸϵ��������һ����2.5��2����ǰ���������O3Ϊ�������Ⱥ������ĸϵ������y-O��Լ��5����ǰ�����ڱ����������࣬Ȼ�����齭ˮϵ��Ǩ�㣬��4��3����ǰO2��O1��O3�Ⱥ�������̨��ԭס��������ĸϵ����Ⱥ�����伸�������˶�����Ⱥ������Nϵ����(��A��N9b)������B4��B5��F��N9a��R9�ȣ������Mϵ��M7b��M7c����˵����O2��O1��O3����֮ǰ��y-O��Ⱥ���Щĸϵ����Ⱥ�Ѿ����������ȵطֲ��ڸ������䣬�����ں����������зֱ���O1��O2��O3���Ӧ�Ĵ�Ķ������͡���������ԭס����D(D4,D5)��G��M8a��Ӧ��Դ��ʢ��������y-D��Ⱥ��ĸϵ������D5����(38.3ka)����Ǩ������磬�ϵ���Ⱥ������D4����(27ka)���ձ���ڶ�����Ⱥ�����ϵ�������mt-C������������������δ�����ϵ���̨�塢����ԭס������y-Oϵ�Ľ������ܽ������ӳ��������ε����������ϻ�����1.5����ǰ��(99) ��������.��.���� ��ʯ��ʱ������Oϵ��ռ���ϣ���ʯ��������Դ�������ϵĸ�ϵ����Ⱥ��C��D��O��N��Oϵ�����Ϸֻ������ţ�ֱ����ʯ�����ڸ�ϵD��C��N��δǨ�����ϡ��ж����ݣ�ʢ���ڴ����Ϸֳ���̨��ԭס��������û��D��C(�����,�IJ�)����û��N��ʢ���ڴ�����Ǩ���ϵ�����O2a-M95Ϊ�������Ⱥû����D��N(��natstuya)����ʯ��ʱ������ʯ�����ڴ�����Ǩ�����ϰ뵺���ϸ�����ȺD����(��������)��N�ັ�У����϶�����ȺD�����ڸ�����Ⱥ��Cϡ�٣������붫Խ�йص���Ⱥ��һ������(�����,�IJ�)������ӷ���Ǩ��ʱ��C�Ӷ������غ���D�Ӳض������ϣ�����C3�عź���Ǩ�㱱����N�����ϵ�����Ǩ������Oϵһֱ��ռ�����ϣ�ֱ����ʯ�����ھ�������Խ��Ǩ����Ȼ��ϵO(O1��O2��O3)���ܱߵ�C��D��N�������ںϣ���ĸϵ���н����ģ�������������(��baiyueren)��̨��ԭס��(��natstuya)��D(D4,D5)��G��M8a��Ӧ����������һ����y-D��Ⱥ����Щĸϵ����ȺҲ�㷺�ֲ��ڶ�����Ⱥ����(���IJ�)��(98) ��������.��.���� ��ʯ��ʱ������y-D��Ⱥ��ŷ�����֧ϵ���������ǰ��y-D��Ⱥ��ӡ�ȶ���Ǩ��ض�������һ������������������Ǩ��ܿ죬�ô�Լ1����ʱ��ͱ鼰�˱�������������D��Ⱥ���������֧ϵ��һ֧ϵ�زش�����Ǩ��������ϵ������D*��D1��������������D3��ԭ����D��G��M8��M9��M10�ȣ������ֵ�����C��Z�ȣ��������������������ε�A��ˮ������Ӧ����һ֧ϵ������������ɷ�Ӧ������һ֧������Ǩ�㱱����y-N��Ⱥ��ϸʯ���Ļ�Ӧ����֧�йأ�������ĸϵҲ����һ֧�н�Ե��ϵ����������ĸϵA��C��D��G��M8��M9������һ֧��ԨԴ��ֻ��֮�����y-N��Ⱥ��������֡�D��Ⱥ����һ��֧ϵ�س������£�Ȼ����һ����Ǩ������������֧ϵ��D��D*��D1��ĸϵ����Mϵ�⣬����B��A��F��M7���Ϸ���Ⱥ�����ĵ����ͣ����ֿ���������y-NOϵ�Ľ�����Դͷ�Ǿ��������塢���ȳ����б������͵�y-D*��D1�ײ����ձ��ض�������Ҳ��������һ֧�������عų������Ŷ�������Ǩ���ձ�������;������ձ�ʱ������y-D2(D2��D1��,����D3��Զ)��Ǩ����������2��1.5����ǰ���Ʋ����ڶ���ĸϵ�ʶ������죬�ֽ���������������������ˮ�������߱��ϵ�̫��ɽ���ֽ�������ΪMϵ(��A)���Զ���Nϵ(��M7)Ϊ����Ҳ��Mϵ����Ӧ�ĸ�ϵ��D��C3���ֽ�������������y-N��Ⱥ������Զ���O3(Oϵ)��Ⱥ��������ƺ�ԭʼ��ĸϵ�и���������������(97) ��������.��.���� �����O3Դ�Իƺ������ε�ǰ������Ⱥ��������ʯ��ʱ������ľߴ����ԵIJ��������Ļ��������в�������Ҥ�Ļ�����(����Ϊ��Դ����)���������ռ�����ԭʼ�������Ʒ���Ϊ�������Ƽ�����δ���գ�����͡��������ɡ���ɫ�����ȡ����μ��������������Ļ������ѳ��������Ƽ���������Ҥ���ռ������ǵ���ǰ��δ�еĸ߶ȡ��ɴ��Ʋ⣬��������ɢ��O3��Ⱥ�����ǰ�����Ļ���(8.2��7kaBP)�Ѿ����������������зֳ���O3a1c2-F238��Լ7ǧ��ǰ�����ں���������Ⱥ����������ԭ�������຺���еı���Ϊ2.81%(33/1174)����F238�ڲ����зdz���Ƶ��˵������O3��Ⱥ�ķֳ�Ӧ����F238����֮ǰ(����������������>>>)����Ǩ��֮ǰ���������뺺�干���ĵ�������Ҫ��M117��ռ29.82%������M134*ռ2.63%���乲�������1����7ǧ��ǰ(���ޱ���)�������������ܿ���Ҳ������Դ��������M117�Ĺ������䡣��Ⱥ������Ǩ������ظ�ԭ������ũҵ������ҵ�������²�����Ⱥ�����7��8ǧ���������(���ޱ���)�������������ǿ��ԴӲ����O3ϵͳȥ��̽�ƺ�������ǰ������ԭʼ�۲���ĵ���Ⱥ�����Ӧ��M117��������Ƿȱ002611��Ҳ���ԴӺ��ع��е�Q1��N����Ⱥȥ�Ʋ�ԭʼ��Ⱥ���ں��������ʯ��������в��常ϵ��M9Ӧ��һ������N*��N1��M45�п�����Q1���ޱ�����������2354����N1*ռ5.06%��Qռ1.23%���������жϣ�N��Q1������ǰ�����Ļ�����μˮ�����Ѿ��ںϵ�����������Ⱥ���У�>>>������ԭʼ����ĸϵ����������y-N��Ⱥ�н�Ե��ϵ��ͬ��Mϵ�������ձ��ض������˼�����ӡ�ڰ�������ϵ��Զ(ӡ�ڰ��������ΪA2��B2��C1��D1������X2a��D2��D3��C4c��D4h3)��(96) ��������.��.���� ����ƽ��ĸϵ��Դ���ֵң����ִ�ѧ��������ѧԺ���߽��������ĶԺ��ϰ���������ַ(Լǰ1250��ǰ1050)��СĹ37��ƽ������DNA���������������ĸϵ������ΪD��G��A��C��Z��M10��M*��B��F��N9A�����Ŵ���ϵ����ͭʱ��������������Ⱥ�����������쿪����Ⱥ��ӽ���YȾɫ��DNA���о����ڽ����У���Yungsiyebu���������Ļ�������ƽ��ĸϵ������D��G��A��C��Z��M10��M*�ܿ������Ժӱ���������һ������Դͷ��ӦΪ����-�������������ӹ���ַ֮����ֵҡ�����ƽ��ĸϵ�Ľ��������Դ�ڳ��ǵش���D��G��C��M10��M*��ĸϵ����Ⱥ������ԭ�����Ҫ����ս�������ֵһ��Ļ�������������ַ���������ļҵ��ϲ㶫���Ļ������ļҵ��²��ֵ��Ļ���ʱ��(Լ3300��ǰ)�ܽӽ�����ˣ�����ƽ���ĸϵ��̫������������ļҵ��²��ֵ�(ʱ�����������̴��̹���3600��3046��ǰ)���ٿ��ǵ���ļҵ��²���С�����Ļ�֮������Լ��������Ļ�ȱʧ����һ���ƶϣ��ļҵ��²�������ƽ���ĸϵӦ��Դ������ǧ��ǰ�������������ϼ�����һ�����ֵ�(���ֵ�����)��Լ��ǧ��ǰ���ֵ��뼽�������Ⱥ�����ںϣ�����ͬʱ���ܺ��ں�ɽ�Ļ���Ӱ�죬������ͭʱ�������ڣ�һ����Ǩ��ɽ�Ա����γ���������ַ���ļҵ��²��Ļ��������Ӽ�������Ⱥ�������Ϸ�չ�γ������Ļ������Ļ����Ʋ�������Ⱥ�ĸ�ϵӦ�и߱�����N*��N1��ֻ�Ǻ����౻���ǣ���ĸϵ����Ⱥ������������������Ϊ�ֽ�ӱ�����ĸϵ�����塣��������Դɽ��˵������˵������˵����ȣ���μ��·�������Ը���(����ϵ)�����������ԭ�ȵ�Ң�����йأ���˵Ң������(ɽ��������)����ع��ƹ�(�ӱ�����)���Ƶۺ�Ǩ��ƽ��(ɽ���ٷ�)������ƽ��ĸϵ���ٷ��������������������������ִ����������࣬�������Ϻᱱ��������ε�������Լ��ִ��Ϸ��˾����Զ�����̸�ϵ�����������M134*��002611�� ��������.��.�� ����������ĸϵ��pre-B(����Ϊ��B)��Qiaomei Fu��Matthias Meyerb��Xing Gao�����������ǰ���������˹�DNA�������ʾ�����������൱�������˼�����ӡ�ڰ���������Ե��ϵ������������Ŵ�����ŷ���������ѷ��룻��������Я���ᰲ����������������(��������)DNA���������ȸõ����ִ��˶�(��198401)�����������ִ���(��������)��ĸϵ����Ⱥ��pre-B����B����ĸ��֧���������Ѿ����(��polyhedron��baiyueren)����4.5����ǰ������������Ust'-Ishim�˹�DNA��ø�ϵ��pre-ON��ĸϵ��pre-B�������˵ĸ�ϵ�п���Ҳ��pre-ON��ĩ�μ���ڣ�pre-ON��C��D������Ⱥ�������鼰�������������������ǣ��������ϰ뵺������ֻ�ǵ���ʢ���ڣ�������Ⱥ�������ˣ�ONϵҲ����ֻʣ���Ϸ�O��N����֧�����>>>119��(94) ��������.��.���� ���������������������ϵ��ִ���ͷ�ǻ�ʯ�����������ʿ��Ŷ��ں������������ɽ��(���ξ���)��������������ϵ�6��4.6����ǰ���ִ���ͷ�ǻ�ʯ����Vietschlinger��>>>��Ϊ�ִ��������߳�����һ֧������-���ϰ뵺��Ȼ��ӱ�����������붫���ṩ�˿���ѧ���ݡ������������ϵĸ�ϵ��C��K*��D��ON-M214(O��Nδ����)��CΪ����������Ⱥ��D��ӡ�ȶ����ս��ض������ϣ�K*��Ҫ�ֲ����ϵ������ܴ����ϰ뵺������Ǩ���ϵ�����ʱ��ƽ��Լ�����ڵ�50��70�ף���ˣ����������½��ַ���п�����NO-M214��Ⱥ����Ϊ����Oϵ������Nϵ�����ȣ���Ӧĸϵ������B*��N*��N9��R9��F��M7�ȡ�(93) ��15�� ��14�� ��13�� ��12�� ��11�� ��10�� ������ ������ ������ ������ ������ ������ ������ ������ ������ |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}